|
|
..... |
PrIBES: Proyecto para Iberoamérica de Entomología
Sistemática. 2000
NEOTRÓPICO:
Sistemática
y filogenia de los himenópteros de la región
Neotropical:
Estado del conocimiento y perspectivas
Fernando
Fernández C.
Instituto
Humboldt
Apartado Aéreo 8693, Santafé de Bogotá
D.C.
COLOMBIA
ffernandez@humboldt.org.co
Resumen:
Hymenoptera
es uno de los órdenes de insectos más diversos
en el mundo. La región Neotropical es muy rica, aunque
la magnitud real de la cantidad de especies se desconoce.
Se presenta una síntesis del conocimiento del grupo,
incluyendo número de familias, subfamilias, tribus,
géneros y especies. Hymenoptera comprende 20
superfamilias, 77 familias, 2.527 géneros y cerca de
24.000 especies en el Neotrópico. Se ofrece una breve
guía de identificación para la mayoría
de los taxones. Probablemente el número real de especies
de himenópteros neotropicales pase de 60.000, muchas
de éstas sin describir, en contraste con el pobre número
de taxónomos dedicados a este grupo.
Palabras
clave: Hymenoptera,
Sinopsis, Región Neotropical.
Systematics
and phylogeny of Neotropical Hymenoptera: State of the art
and prospects
Abstract:
The Hymenoptera
is one of the most diverse insect orders in the world. The
Neotropical region is very rich, although the real magnitude
of its species richness is unknown. A synthesis of current
knowledge about of the group is presented, including numbers
of families, subfamilies, tribes, genera, and species. The
Hymenoptera comprises 20 superfamilies, 77 families,
2,527 genera and about 24,000 described species in the Neotropics.
A short guide to their identification and a list of many taxa
are presented. The real size of the Neotropical hymenopterofauna
may surpass 60,000 species, most of them undescribed, which
contrasts with the poor number of taxonomists devoted to their
study.
Key
words: Hymenoptera,
Synopsis, Neotropical Region.
Introducción
Se
considera que las avispas, abejas y hormigas comprenden uno
de los grupos más diversos del reino animal, el orden
Hymenoptera (Nieves-Aldrey y Fontal-Cazalla, 1999). Dentro
de los insectos, Coleoptera, Hymenoptera, Lepidoptera
y Diptera constituyen los cuatro órdenes
hiperdiversos, comprendiendo en conjunto la gran mayoría
de los seres vivos en el planeta (Gaston, 1993). En el caso
de los himenópteros, se han descrito alrededor de 120.000
especies (Borror et al., 1989; Minelli, 1993) y se estiman
más de 200.000 (Gauld y La Salle, 1993).
La importancia de los himenópteros radica no sólo
en su número, sino en sus funciones y adaptaciones
ecológicas. La vasta mayoría de especies son
parasitoides, atacando gran cantidad de artrópodos,
especialmente otros insectos. Muchas especies de importancia
ecológica y económica están reguladas
por estas avispas. La polinización de muchas plantas
con flores depende de las abejas y otros himenópteros.
Las hormigas son insectos dominantes en los ecosistemas tropicales,
donde influencian la estructura y dinámica de suelos
y árboles. Dentro del orden ha surgido la sociabilidad
varias veces, además de varias formas de subsociabilidad
y parasitismo (Gauld y Bolton, 1988). Algunos grupos de himenópteros
se utilizan o proponen en estudios biogeográficos y
de conservación y monitoreo (p.e. Brown, 1991; Perfecto
et al., 1997).
Estudios en diferentes campos de la biología reconocen
a los himenópteros como un grupo monofilético
y de larga historia, con fósiles conocidos desde el
Triásico (Rasnytsin, 1988). La clasificación
superior del grupo (subórdenes y superfamilias), durante
largo tiempo poco modificada, está sufriendo importantes
cambios, gracias sobre todo a la influencia de la cladística
y la biología molecular. No existen acuerdos firmes
sobre la clasificación de las categorías superiores
y el estatus de muchos grupos importantes es aún incierto
(Whitfield, 1998).
Familias de gran importancia como hormigas, abejas, esfécidos
y avispas papeleras se conocen relativamente bien desde el
punto de vista sistemático. Sin embargo, son muchas
las familias con un conocimiento muy pobre (como muchos grupos
de microhimenópteros) en relación inversa con
su riqueza e importancia. El panorama se complica ante la
gradual e imparable 'extinción' de taxónomos
de estos insectos, únicas personas que podrían
ofrecer bases sólidas para estudios de otra naturaleza.
En este documento se ofrece un vistazo al conocimiento en
sistemática y biología de los himenópteros,
con énfasis en la región Neotropical.
Monofilia
de Hymenoptera
Desde
hace mucho tiempo se considera que avispas, abejas y hormigas
comprenden un grupo separado de los demás insectos;
el mismo nombre del orden alude a los hámulos que permiten
unir las alas anteriores con las posteriores para un vuelo
sincronizado (Goulet y Huber, 1993). Este carácter
es autapomórfico para el orden, y prueba de su monofilia,
pues no se conoce en otros insectos (Königsman, 1978).
Secundariamente se han propuesto otros atributos basados en
la estructura del aparato bucal, venación y genitales
(Königsman, 1978). En los últimos años
la morfología externa comparada, anatomía interna,
paleontología y reinterpretación de caracteres
han arrojado un buen número de caracteres como potenciales
apomorfías para el orden. En la Tabla
I se relacionan estos atributos, basados en una reciente
recopilación de Vilhelmsen (1997b). La haplodiploidía
no se ha verificado en algunos grupos menores y es convergente
pues se conoce en otros invertebrados (Wilson, 1971; Crozier,
1977).
Estudios
de morfología y biología en Hymenoptera
Hymenoptera
es un orden extensamente estudiado en varios aspectos de su
biología. Algunos aspectos de interés son: morfología
del limpiador de antena y placas sensibles (Basibuyuk y Quicke,
1995, 1999), cabeza (Vihelmsen, 1997a), mesosoma y músculos
asociados (Gibson, 1985, 1993; Whitfield et al., 1989; Heraty
et al., 1994), espiráculos (Tonapi, 1958a,b,c,d), mesocoxas
(Johnson, 1988), ovipositor (Quicke et al., 1992a, 1994),
esperma (Quicke et al., 1992b), formas inmaduras (Evans, 1987).
Godfray (1994) y Quicke (1998) revisan la extensa literatura
en biología de parasitoides. Stolz y Whitfield (1992)
revisan el interesante tema de virus y entidades viroidales
asociadas. Quicke et al. (1999b, c) presentan recomendaciones
para preservación de especímenes para estudios
en morfología y biología molecular. Eady (1968),
Harris (1979), Gauld y Bolton (1988), Goulet y Huber (1993)
y Hanson y Gauld (1995) ofrecen términos, definiciones,
glosarios o ilustraciones útiles para la morfología
y biología básicas en el orden. La Salle y Gauld
(1991, 1993) presentan diversos acercamientos al estudio de
los himenópteros en la perspectiva de la diversidad
biológica y su conservación. Godfray (1994)
y Quicke (1997) ofrecen revisiones en la biología de
parasitoides, Hölldobler y Wilson (1990) en biología
de hormigas y Roubik (1989) en historia natural de las abejas
de los trópicos.
Ubicación de Hymenoptera dentro de Insecta
Los
insectos son un grupo monofilético (Kristensen, 1991)
dentro de los artrópodos. La filogenia interna de la
clase dista de ser estable, aunque se acepta que los insectos
holometábolos conforman un grupo monofilético
(Kristensen, 1991), gracias a la existencia de formas inmaduras
(larvas) muy distintas de los adultos. La figura 1 muestra
una propuesta filogenética de la superclase Hexapoda,
de acuerdo con las ideas de Kristensen (1991).
Dentro de Endopterygota (Holometabola) se reconocen
dos grandes linajes, el neuróptero-coleopteroide y
el himenóptero-mecopteroide; el primero de estos comprende
órdenes como coleópteros y rafidiópteros
y el segundo himenópteros, mecópteros, pulgas,
moscas, mariposas y trips. En el grupo de los himenóptero-mecopteroides
Hymenoptera aparece basalmente, como el grupo hermano
de Mecopterida (Mecoptera a Lepidoptera,
Fig. 1).
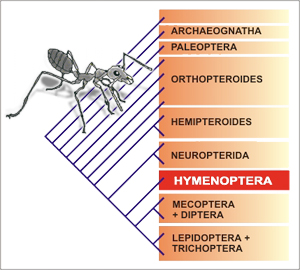
Fig.
1.
Filogenia de las grandes divisiones de Hexapoda, mostrando
la
posición de Hymenoptera (basado en Kristensen, 1991)
Sistemática
de los grandes grupos, con énfasis en la región
Neotropical
La
sistemática de nivel suprafamiliar está lejos
de la estabilidad. Durante mucho tiempo se aceptaron tres
grandes grupos: “Symphyta” (moscas sierra),
“Parasitica” (avispas parasitoides) y avispas
con aguijón (“Aculeata”). Sin embargo,
de estos tres grupos, al parecer solamente el último
es monofilético (Brothers, 1975; Brothers y Carpenter,
1993), y los otros son parafiléticos. Recientes estudios
filogenéticos proponen arreglos diferentes (Brothers
y Carpenter, 1993; Vihelmsen, 1997b; Whitfield, 1998) con
un sistema de relaciones genealógicas internas diferente
a los tradicionales (Fig. 2).
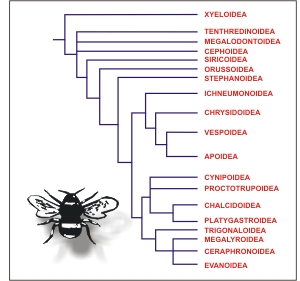
Fig.
2. Filogenia de las superfamilias de Hymenoptera
(basado en Whitfield, 1998)
Las superfamilias presentan un poco más de estabilidad,
y aun más las familias, donde en varios casos las disputas
son sobre el nivel de jerarquía y no sobre su estado
natural (p.e. una sola familia, Apidae, o varias, para
agrupar todas las abejas). El descubrimiento de nuevos caracteres
(especialmente internos), la reinterpretación de algunos
caracteres externos, y el uso de técnicas moleculares
parecen abrir nuevas perspectivas en el estudio del origen,
evolución, filogenia y clasificación de las
familias de Hymenoptera.
Ronquist (1999a) ofrece una síntesis actualizada sobre
el conocimiento actual en filogenia de Hymenoptera;
Dowton y Austin (1994) estudian las relaciones en Apocrita
a partir de una filogenia molecular; Ronquist et al. (1999)
proponen un análisis filogenético del orden
basados en Rasnytsin (1988); Vilhelmsen (1997b, 1999) y Dowton
y Austin (1999) revisan la filogenia de los himenópteros
más basales (los antiguos “Symphyta”);
Whitfield (1992, 1998) realiza una síntesis de estudios
en filogenia del vasto conjunto de avispas parasitoideas;
Sharkey y Wahl (1992) estudian las relaciones filogenéticas
en Ichneumonoidea, y Belshaw et al. (1998), Quicke
et al. (1999a) exploran aspectos paleontológicos, morfológicos
y moleculares en Ichneumonoidea; Achterberg y Quicke
(1992) reinterpretan la filogenia de las subfamilias de Braconidae.
Dowton et al. (1997) estudian la filogenia molecular de proctotrupomorfos
y evaniomorfos; Alexander (1992), Brothers y Carpenter (1993),
Brothers (1999) y Carpenter (1999) revisan la filogenia de
los himenópteros con aguijón (Chrysidoidea,
Apoidea, Vespoidea). Gibson (1986, 1999) y Gibson,
Heraty y Woolley (1999) exploran la filogenia de Chalcidoidea,
Mymarommatoidea y Platygastroidea. Ronquist
(1995, 1999b) estudia la clasificación, evolución
y filogenia en Cynipoidea y Nordlander et al. (1996)
exploran las relaciones de Ibaliidae. Shaw (1990) presenta
la filogenia y biogeografía de la enigmática
familia Megalyiridae. Gauld y Bolton (1988) ofrecen
un buen panorama a la morfología y biología
del orden, y Goulet y Huber (1993) editarn un manual de identificación
de las familias del mundo, con riqueza de ilustraciones y
referencias bibliográficas. En fin, Carpenter y Wheeler
(1999) discuten los análisis simultáneos morfológicos
y moleculares en el orden.
La región Neotropical posee una fauna grande y rica
en formas autóctonas de himenópteros (Hanson
y Gauld, 1995) pero sabemos muy poco de su historia, biología
y sistemática. Tampoco se sabe cuantas especies se
han descrito, ni se han realizado estimaciones. Costa Rica
ha sido objeto de intensos muestreos de su himenopterofauna
(Hanson y Gauld, 1995), que se estima en 17.000 especies (Gaston
et al., 1996). Ello implica una gran diversidad para el país,
pues con un área muy pequeña tendría
prácticamente el mismo número de especies que
toda la región Neártica (Canadá, EE.UU.
y norte de México, con 16.500 especies conocidas).
Aun así, muchos grupos de himenópteros son pobre
o nulamente conocidos en este país (Gaston et al.,
1996). La información procedente de este país
sugiere que la región Neotropical debe poseer un número
enorme de himenópteros, del orden de 70.000 especies,
con al menos un 80% sin describir. Como ejemplo, Townes y
Townes (1966) catalogan unas 1800 especies de Ichneumonidae
para el Neotrópico y estiman en 20.000 las especies
para esta región.
Hasta la fecha no existen manuales de identificación
para la fauna neotropical. El libro editado por Hanson y Gauld
(1995) es una buena herramienta para la fauna mesoamericana
(y un poco para la fauna del norte de Suramérica),
pero de poca ayuda para la fauna de zonas frías y caribeña.
Las familias que poseen claves al menos hasta género
y que cubren toda la región Neotropical son Argidae,
Pergidae, Xiphydriidae, Orussidae, Braconidae,
Evaniidae, Ceraphronidae, Megaspilidae,
Cynipidae, Ibaliidae, Liopteridae, Eucolilidae,
Megalyridae, Leucospididae, Chalcididae,
Encyrtidae, Mymaridae, Heloridae, Platygastridae,
Proctotrupidae, Scelionidae, Bethylidae,
Dryinidae, Chrysididae, Rhopalosomatidae,
Scoliidae, Tiphiidae y Vespidae.
En Goulet y Huber (1993) y Hanson y Gauld (1995) se encuentran
las referencias bibliográficas para la identificación
de los géneros de estas familias, o se proponen claves
para géneros de algunas familias de tamaño pequeño.
Las siguientes familias, que no están relacionadas
en esas dos publicaciones, en lo referente a modernas claves
para géneros, son: Stephanidae (Aguiar, 1998),
Plumariidae (Roig-Alsina, 1994), Embolemidae
(Olmi, 1995b), Scolebythyidae (Azevedo, 1999), Sphecidae
(Menke y Fernández, 1996), Apidae (Michener
et al., 1994; Michener, 2000), Formicidae (Bolton,
1994), Braconidae (Wharton et al., 1997) y Trigonalidae
(Carmean y Kimsey, 1998).
Existen catálogos para la mayoría de grupos
de himenópteros que cubren total o parcialmente la
región Neotropical. La mayoría de estos son
del siglo pasado o comienzos de éste, por lo cual no
sólo no están actualizados sino que además
generalmente son inaccesibles. Catálogos recientes
que cubran la región Neotropical son los de “Symphyta”
(Smith, 1978), Trigonalidae (Weinstein y Austin, 1991),
Chalcidoidea (De Santis, 1979, 1983, 1989; De Santis
y Fidalgo, 1994), Proctotrupoidea (Jonhson, 1992),
Platygastridae (Vlug, 1995), Ichneumonidae (Townes
y Townes, 1966), Chrysididae (Kimsey y Bohart, 1990),
‘Sphecidae’ (Bohart y Menke, 1976), Formicidae
(Kempf, 1972; Brandao, 1991; Bolton, 1995) y Mutillidae
(Nonvellier, 1990). En superfamilias importantes como Cynipoidea
o Evanioidea no existen catálogos para la región
Neotropical. Pagliano y Scaramozzino (1990) ofrecen el listado
sinonímico de los 17.000 géneros y subgéneros
de himenópteros fósiles y vivientes del mundo.
Xyeloidea
Considerado
el grupo más basal dentro de Hymenoptera, conocida
desde el Triásico medio, hace más de 200 m.
a. (Rasnytsin, 1988). Especies fitófagas. Vihelmsen
(1997a) estudia la monofilia y relaciones del grupo. Una sola
familia, Xyelidae, holártica con sólo
un género y una especie en la región Neotropical
(Smith, 1988).
Tenthredinoidea
La
superfamilia más numerosa de los tradicionales “Symphyta”.
Aunque se considera monofilética, las relaciones internas
y la monofilia de las familias son inciertas (Vihelmsen, 1997b,
1999). Grupo conocido desde el Cretáceo, con seis familias
y alrededor de 7.000 especies vivientes en el mundo. Todas
sus formas recurren a fuentes de alimentación de origen
vegetal (Smith, 1993). Argidae es una familia bien representada
en los trópicos, con alrededor de 400 especies, revisadas
por Smith (1992). Cimbicidae es una familia de zonas
templada con menos de 10 especies neotropicales (Smith, 1988).
Diprionidae, taxón de climas fríos (donde
es plaga de algunas plantas), con menos de 15 especies neotropicales
(Smith, 1988). Como Argidae, Pergidae es un
taxón importante en los trópicos, con alrededor
de 300 especies neotropicales (Smith, 1990). Tenthredinidae
es la familia más grande y común en la región
Neotropical, con alrededor de 400 especies (Smith, com. pers.).
Megalodontoidea
Superfamilia monofilética, principalmente holártica,
con sólo cuatro especies neotropicales, en la familia
Pamphilidae (Smith, 1988).
Cephoidea
Superfamilia monotípica y holártica, con sólo
una especie que coloniza la región Neotropical (Smith,
1988).
Siricoidea
Probablemente es esta una superfamilia polifilética,
con diversos puntos de vista sobre sus límites y relaciones
internas (Gauld y Hanson, 1995; Vihelmsen, 1997b). Es un grupo
principalmente holártico, con una decena de especies
en la familia Siricidae y 17 en Xiphydriidae
(Smith, 1988). Para algunos autores esta última familia
es de posición incierta (Goulet y Huber, 1993).
Orussoidea
Superfamilia monofilética y monotípica. Se considera
el grupo hermano de Apocrita (Whitfield, 1998). Probablemente
en el ancestro común de Orussoidea y Apocrita
surgió la depredación o el parasitoidismo,
partiendo de ancestros fitófagos. Este hecho, sucedido
quizás en la transición entre el Triásico
y Jurásico, es el responsable de la enorme expansión
de especies y formas de vida desde entonces y sobre todo en
el Terciario. Aparentemente estos himenópteros fueron
ectoparásitos de insectos de la madera, comportamiento
que se considera generalizado en Apocrita (Gauld y
Hanson, 1995). Orussidae es una familia cuyas larvas
atacan coleópteros y otros himenópteros (Smith,
1993). Como Xiphydriidae, es una de las familias más
raras del orden en los trópicos. Se conocen 13 especies
neotropicales, revisadas en Smith (1988).
Stephanoidea
Superfamilia monofilética y monotípica. Se considera
la familia más basal dentro de Apocrita. Ectoparásitos
de coleópteros de la madera. Grupo aparentemente más
común en épocas pasadas, actualmente más
conocida del hemisferio sur. La vieja revisión de Elliot
(1922) registra menos de 30 especies, número que puede
ser inferior al real, pues Aguiar (1998) lista 26 especies
en el género Hemistephanus para Suramérica.
Este autor reconoce tres géneros neotropicales.
Trigonalioidea
Superfamilia
monotípica. Por su biología y morfología
es uno de los grupos más enigmáticos del orden;
no hay acuerdo sobre su posición dentro de Hymenoptera
(Whitfield, 1998). Presentan diversos grados de parasitismo
y huéspedes primarios y secundarios (Weinstein y Austin,
1991). Carmean y Kimsey (1998) estudian la filogenia interna
del grupo y presentan claves para los géneros. Weinstein
y Austin (1991) revisan la biología y ofrecen el catálogo
de las especies descritas.
Ichneumonoidea
Se
acepta actualmente que Ichneumonoidea y Aculeata
(Vespomorpha) son grupos hermanos, conformando a su
vez un grupo monofilético dentro de Hymenoptera
(Rasnytsin, 1988; Sharkey y Wahl, 1992; Whitfield, 1998; Quicke
et al., 1999a). A su vez, Sharkey y Wahl (1992) presentan
la monofilia de Ichneumonoidea y reducen el número
de familias a dos, Braconidae e Ichneumonidae.
Este es un taxón enorme, con alrededor de 100.000 especies
estimadas en el mundo. Las avispas en este grupo presentan
gran variación en formas de parasitismo, con ectoparásitos,
endoparásitos, idiobiontes y cenobiontes (Godfray,
1994). Igualmente, se conoce el hecho de asociación
con virus para debilitar o eliminar las defensas de sus víctimas
(Whitfield, 1992; Stoltz y Whitfield, 1992).
Ichneumonidae es una familia monofilética,
con una cifra estimada de 60.000 especies (Sharkey y Wahl,
1992; Gauld, en Hanson y Gauld, 1995). A pesar de su tamaño,
ha sido objeto de una serie de revisiones por parte de Townes
(1969, 1970a, b) quien caracteriza la mayoría de los
géneros. Wahl (en Goulet y Huber, 1993) presenta claves
para las subfamilias. Townes y Townes (1966) ofrecen un catálogo
para las especies neotropicales, y Yu y Horsmann (1997a, b)
ofrecen el catálogo más actualizado para todo
el mundo, incluyendo museos depositarios. Se han descrito
420 géneros y alrededor de 3137 especies neotropicales
(Palacio, com. pers.).
Braconidae, con alrededor de 40000 especies estimadas
para el mundo, ocupa el segundo lugar en número de
especies. Familia monofilética, con hábitos
tan diversos como Ichneumonidae, y de gran importancia económica.
Wharton et al. (1997) ofrecen una guía de identificación
(también publicada en español) para las subfamilias
y géneros de América. Se han descrito alrededor
de 1.500 especies neotropicales (Sharkey, com. pers.).
Vespomorpha
(=Aculeata)
Este
es un grupo monofilético, compuesto por tres superfamilias
monofiléticas (Chrysidoidea, Apoidea y
Vespoidea). Brothers (1975), Brothers y Carpenter (1993)
y Brothers (1999) estudian la filogenia de Aculeata.
Chrysidoidea
La
superfamilia más basal de Aculeata. Filogenia
estudiada por Carpenter (1986a, 1999) y Brother y Carpenter
(1993). Comprende siete familias monofiléticas con
algo más de 1000 especies neotropicales. Plumariidae
es el grupo basal dentro de la superfamilia, con distribución
gondwánica y alrededor de 18 especies en el Neotrópico.
Claves en Roig-Alsina (1994). Scolebythidae es una pequeña
familia descrita para el neotrópico y Australia con
dos géneros y tres especies en el Neotrópico
(Evans, 1963; Azevedo, 1999). Bethylidae es la familia
más grande de la superfamilia, con más de 3.000
especies en el Mundo. Algunas especies son ectoparásitas
de coleópteros, microlepidópteros y avispas
esfécidas. Se conocen menos de 500 especies en la región
Neotropical (Evans, 1964).
La segunda familia en número de especies es Chrysididae
(“avispas cuclillo”). Se trata de especies ectoparásitas
de “sínfitos”, fasmópteros y de
himenópteros nidificadores. Se conocen alrededor de
280 especies neotropicales; Kimsey y Bohart (1990) ofrecen
una síntesis de biología, sistemática
y listado de especies para todo el mundo. Scleroggibidae
es una pequeña familia, muy raramente coleccionada
en la región. Hasta donde se sabe, son parasitoides
de Embiidina, con apenas dos especies neotropicales
(Richards, 1939; Argaman, 1988; Fernández et al., 1996).
Dryinidae es una familia que se ha especializado en
atacar homópteros auquenorrincos (Cicadelloidea,
Fulgoroidea y algunos Membracoidea), con formas
endoparásitas y ectoparásitas. Se han descrito
alrededor de 430 especies neotropicales, cuya biología
se revisa en Olmi (en Hanson y Gauld, 1995).
Las especies del mundo han sido revisadas en Olmi (1984, 1986,
1989, 1993a, b, 1995a). Guglielmino y Olmi (1997) ofrecen
el catálogo de drínidos y sus huéspedes
en el Mundo. Embolemidae es una familia rara, de biología
desconocida, con cinco especies neotropicales revisadas por
Olmi (1995b).
Apoidea
Esta
superfamilia es el grupo hermano de Vespoidea (Brothers
y Carpenter, 1993). Grupo monofilético, cuyas relaciones
internas explora Alexander (1992). Comprende dos divisiones,
esfeciformes y apiformes. Se considera que las abejas son
“esfécidos modificados” (Gauld y Hanson,
1995). Los esfeciformes se han divido en una sola familia
(Bohart y Menke, 1976) o varias (Goulet y Huber, 1993). Recientemente
se establecieron tres familias: Ampulicidae [grupo
hermano de Sphecidae s. str. + (Crabronidae
+ Apiformes)], Sphecidae s. str. y Crabronidae
(Brothers, 1999; Melo, 1999). Por esta razón el término
"esfeciformes" se puede utilizar como un término
informal para estas tres familias o para todos aquellos apoideos
que no son abejas.
Los
esfeciformes comprenden gran variedad de formas solitarias,
comunales y un género eusocial con hábitos variados
de caza y nidificación. Alrededor de 1.700 especies
habitan el neotrópico. Se han publicado claves de géneros
(en español e inglés) y literatura de identificación
por Menke y Fernández (1996). Los apiformes se han
divido en una familia (Gauld y Bolton, 1988) o varias (Goulet
y Huber, 1993). Roig-Alsina y Michener (1993) y Alexander
y Michener (1995) exploran la filogenia de las abejas de lengua
larga y corta, respectivamente.
Las conclusiones de estos autores se siguen aquí, aceptando
5 familias para la región Neotropical: Colletidae
(442 especies), Andrenidae (incluyendo Oxaeidae,
238 especies), Halictidae (alrededor de 800 especies),
Megachilidae (incluyendo Fideliidae, alrededor
de 350 especies) y Apidae (incluyendo Anthophoridae,
alrededor de 1500 especies). Michener (2000) ha estudiado
la biología, filogenia y sistemática de todas
las abejas del Mundo incluyendo claves para todas las categorías
supraespecíficas. Michener et al. (1994) ofrecen claves
y guía de identificación (en inglés y
español) para las abejas de Norte y Centroamérica
y Moure y Hurd (1987), el catálogo de los Halictidae
del Hemisferio Occidental.
Vespoidea
A
pesar de su tamaño e importancia, no hay sólidas
autapomorfias para Vespoidea (Brothers y Carpenter,
1993), lo cual no impide considerarlo un grupo monofilético.
Grupo con gran riqueza en tipos de vida y biología,
con formas eusociales (biología en Gauld y Bolton,
1988; Hanson y Gauld, 1995). Comprende 10 familias y alrededor
de 6000 especies neotropicales. Sierolomorphidae es
el grupo basal en la superfamilia (biología desconocida)
con unas pocas especies en Mesoamérica (Evans,1963).
Rhopalosomatidae comprende formas parasitoides de grillos
con alrededor de 20 especies en la región Neotropical
(Townes, 1977a).
Bradynobaenidae es un grupo muy poco estudiado, con
menos de 15 especies descritas (Genise, 1986; Quintero y Rodríguez,
1990). Las hormigas comprenden un grupo monofilético
con 17 subfamilias cuyas relaciones internas estudian Baroni
et al. (1992) y Grimaldi et al. (1997); las casi 3000 especies
neotropicales están en el catálogo de Kempf
(1972) actualizado por Brandao (1991). Catálogo mundial
en Bolton (1995) y claves para los géneros del mundo
en Bolton (1994). Hölldobler y Wilson (1990) ofrecen
una síntesis de la biología del grupo.
Vespidae comprende a las avispas “alfareras”,
las eumeninas, y las “papeleras” con formas sociales.
Familia monofilética, así como cada una de sus
subfamilias (Carpenter, 1982). Se han publicado claves para
géneros de Masarinae en Richards (1962) y Carpenter
(1989); Eumeninae (Carpenter, en prep.) y Polistinae
(Richards, 1978). Los Vespidae reúnen alrededor
de 1400 especies neotropicales. Scoliidae es una pequeña
familia de avispas que atacan larvas de lamelicornios, con
una veintena de especies neotropicales (Bradley, 1945). Tiphiidae
comprende avispas ectoparásitas de insectos del suelo.
Kimsey (1991) estudia la filogenia de las subfamilias.
La subfamilia más grande en el Neotrópico es
Thynninae, de la cual se desconoce el número
de especies (Kimsey, 1992). Las avispas cazadoras de arañas
constituyen la familia monofilética Pompilidae,
grupo con alrededor de 800 especies en la región. Shimizu
(1994) explora la filogenia interna en la familia. A pesar
de algunos trabajos básicos (Banks, 1946, 1947; Evans,
1966) esta familia está muy mal estudiada en Suramérica.
Mutillidae comprende las “avispas afelpadas”
con alrededor de 1.500 especies neotropicales (Nonvellier,
1990). Hembras muy conspicuas y bien armadas para el ataque
a nidos de otros himenópteros. No existen claves para
los géneros de la región. La familia Sapygidae,
avispas parasitoides de otros himenópteros, es muy
pobremente conocida (Pate, 1947).
Cynipoidea
Aunque
esta superfamilia es monofilética, sus relaciones internas
y ubicación dentro de la filogenia de Hymenoptera
constituyen un punto de variadas interpretaciones. Comprende
formas parasitoides, formadoras de agallas e inquilinas (Hanson
y Gauld, 1995). Las propuestas de clasificación de
altas categorías en la familia son objeto de puntos
opuestos, de acuerdo a las propuestas de Fergusson (1995)
y Ronquist (1995a, 1999b). Aquí se sigue provisionalmente
el esquema de este último autor. Liopteridae
es un grupo relativamente basal que ataca insectos de la madera,
con unas 60 especies neotropicales (Ronquist, 1995b). Figitidae
es una familia grande, con parasitoides de dípteros
y otros grupos, con más de 300 especies descritas (Weld,
1952). Esta familia comprende Eucoilini, una tribu
con alta diversidad de géneros y especies en la región,
muy pobremente estudiados. En la familia Cynipidae
se encuentran las formas agallíferas e inquilinas,
aunque este grupo es más común hacia las regiones
frías anfitropicales. Cynipodea es uno de los
grupos menos conocidos en los trópicos y hasta la fecha
la única fuente de información es Weld (1952).
Isidoro et al. (1999) estudian la diversidad y función
de las glándulas antenales en machos de esta superfamilia.
Proctotrupoidea
Como
señala Masner (en Hanson y Gauld, 1995) éste
es un grupo polifilético o parafilético que
se ha usado para acomodar varios grupos heterogéneos
de avispas parasitoides (Naumann y Masner, 1985). Las familias
en que se dividen podrían tener poco que ver entre
sí. De éstas, seis se registran en la región
Neotrópica. Monomachidae (10 especies, parasitoides
de moscas), Heloridae (dos especies, endoparásitas
de Chrysopidae) y Pelecinidae (una especie,
ataca larvas de lamelicornia) son pequeñas familias
de biología y sistemática poco conocidas. Proctotrupidae
(100 especies) comprende formas endoparásitas cenobiontes
de larvas de Coleoptera y Diptera; han sido
revisadas por Townes y Townes (1981). Diapriidae es
la familia más grande de esta superfamilia, con más
de 200 especies neotropicales.
Además de las especies endoparásitas de otros
insectos, existen numerosas formas asociadas a hormigas, algunas
miméticas de estos insectos. Johnson (1992) presenta
el catálogo de los Proctotrupoidea s.l. del
mundo, excepto Platygastridae.
Platygastroidea
Esta
superfamilia, aparentemente monofilética, fue extraída
de Proctotrupoidea (Masner en Hanson y Gauld, 1995)
y comprende actualmente sólo dos familias, cada una
monofilética. Para Gibson (1986, 1999) Platygastroidea
es grupo hermano de Chalcidoidea + Mymarommatoidea.
Masner (1976b, 1980) ofrece claves de los géneros de
la familia Scelionidae (334 spp), endoparásita
de huevos de insectos. Masner y Huggert (1989) tratan la mayoría
de géneros de Platygastridae (alrededor de 120
especies), familia de endoparásitos de huevos y formas
inmaduras de otros insectos. Vlug (1995) ofrece el catálogo
de Platygastridae del Mundo.
Mymarommatoidea
Superfamilia
monotípica de biología desconocida. Un género
y unas pocas especies descritas, de morfología enigmática
(Goulet y Huber, 1993). Se considera el grupo hermano de Chalcidoidea
(Gibson, 1986; Gibson et al., 1999).
Chalcidoidea
Superfamilia
monofilética. Grupo muy grande de avispas (la mayoría
de pequeño tamaño) con alrededor de 20.000 especies
en todo el Mundo. Se han propuesto muchos arreglos de la sistemática
de familias en el grupo, lo cual ha creado numerosas clasificaciones
(Boucek, 1988). Goulet y Huber (1993) presentan una propuesta
intermedia con 21 familias, aunque falta más estudio
para definir claramente los límites de las familias.
Se acepta que varias familias son artificiales (como Pteromalidae)
y probablemente el número real de éstas sea
inferior al aceptado actualmente.
Tan compleja como su filogenia es su biología con un
amplio espectro de formas de vida, con formas idio y cenobiontes,
endo y ectoparásitas, atacando muchos grupos de insectos
y arácnidos y con parasitismo, hiperparasitismo y poliembrionía
(Godfray, 1994; Quicke, 1998). Algunos son parasitoides especializados
(como los Eucharitidae que atacan hormigas), otros son obligados
polinizadores (como las Agaonidae en Moráceas),
otros hacen agallas (como algunas Pteromalidae), o
atacan huevos (Mymaridae), aunque la gran mayoría
son parasitoides. Algunos grupos se han revisado y existen
claves por lo menos hasta género (Chalcididae,
Leucospidae, Encyrtidae, Ormyridae, Elasmidae,
Eupelmidae, Trichogrammatidae, Mymaridae).
Megalyroidea
Superfamilia
monotípica y monofilética. Esta es una familia
de distribución “gondwánica” cuyas
relaciones filogenéticas con el resto de Hymenoptera
no están claras (Shaw, 1990). Se sabe que atacan coleópteros
e himenópteros de la familia Sphecidae, y están
representados por tres géneros monotípicos en
Chile y Brasil (Shaw, 1990).
Ceraphronoidea
Superfamilia
monofilética dividida en dos familias. Como Platygastroidea,
estas dos familias formaban parte de Proctotrupoidea
s.l. hasta que se separaron como grupo independiente (Masner
y Dessart, 1967). Sus especies atacan diversos órdenes
de insectos. Dessart y Cancemi (1986) ofrecen una sinopsis
y claves de los géneros de Megaspilidae y Ceraphronidae.
Evanioidea
En
esta superfamilia no hay seguridad sobre su monofilia, ni
están claras las relaciones entre sus tres familias
componentes (Dowton et al., 1997). Los Aulacidae son
endoparasitoides cenobiontes de insectos de la madera, con
unas 50 especies neotropicales, actualmente en revisión
por D. Smith. Gasteruptiidae comprende cleptoparásitos
de nidos de otros himenópteros, con alrededor de 30
especies en la región Neotropical. Evaniidae
es una familia depredadora de ootecas de cucarachas, con alrededor
de 100 especies.
Síntesis
y perspectivas
Filogenia
Hymenoptera
es un orden claramente monofilético, definido por numerosas
apomorfías en el adulto y por lo menos una en la larva
(Tabla I). La haplodiploidía
es prácticamente universal en el orden, aunque ésta
también existe en algunos coleópteros y ácaros.
Los himenópteros son un grupo antiguo, aparentemente
el grupo hermano de los mecopteroides (Fig.
1). Estudios paleontológicos, morfológicos
y moleculares han permitido ofrecer una propuesta de filogenia
interna en el grupo que, aunque aún insatisfactoria
en algunos puntos, ofrece un panorama cohesionado (Fig.
2).
Además
de una serie de superfamilias basales, se acepta un gran grupo
monofilético en Hymenoptera, los Apocrita. Este gran
grupo a su vez comprende dos grandes divisiones, la primera
formada por Ichneumonoidea más Vespoidea, y
la segunda con el resto de superfamilias parasíticas.
Los estudios muestran que la mayoría de superfamilias
en el orden son monofiléticas. Evidencias claras de
grupos monofiléticos se conocen en las superfamilias
Tenthredinoidea, Megalodontoidea, Cephoidea,
Orussoidea, Stephanoidea, Trigonaloidea,
Ichneumonoidea, Chrysidoidea, Apoidea,
Vespoidea, Cynipoidea, Platygastroidea,
Mymarommatoidea, Chalcidoidea, Ceraphronoidea
y Megalyroidea. Grupos en los cuales su monofilia está
en duda, o son ensamblajes artificiales: Xyeloidea,
Siricoidea, Proctotrupoidea s. str. y Evanioidea.
Los grupos con ubicación problemática dentro
de la filogenia de Hymenoptera son: Cynipoidea, Trigonaloidea
y Evanioidea. Las relaciones de Proctotrupoidea
y Evanioidea con sus vecinos también requieren
más estudios.
Con la excepción de las superfamilias monotípicas,
la mayoría de superfamilias en Hymenoptera aguardan
más estudios para esclarecer sus relaciones internas
y la monofilia de sus componentes. Se conoce la filogenia
de las familias de Ichneumonoidea y las superfamilias
de Aculeata (Vespomorpha) y la mayoría
de las familias se consideran monofiléticas; la única
gran familia parafilética es Sphecidae. En Cynipoidea
se han hecho propuestas de clasificación opuestas,
y deben hacerse otros estudios o reexaminarse los existentes
para llegar a una clasificación estable. Proctotrupoidea
es un ensamblaje artificial, y es urgente un extenso estudio
de sus divisiones para esclarecer sus relaciones. Chalcidoidea
es un grupo muy complejo, y la mayoría de sus familias
están mal definidas. Aunque Evanioidea comprende
tres familias, sus relaciones no están del todo claras,
especialmente la posición de Evaniidae.
Tamaño de Hymenoptera en el Neotrópico
El
orden Hymenoptera comprende 20 superfamilias, 77 familias,
2.527 géneros y alrededor de 24.000 especies descritas
para la región Neotropical. En el Neotrópico
no se han registrado las familias Megalodontidae, Blasticotomidae,
Anaxyelidae, Heterogynaidae, Ibaliidae (sensu
Goulet y Huber, 1993), Austroniidae, Peradeniidae,
Roproniidae, Renyxidae y Vanhornidae;
la mayoría de estas familias son pequeñas o
relictas. No existe acuerdo sobre el número total de
familias en el Mundo; asumiendo el número promedio
de 84 familias, la región Neotropical alberga el 92%
de las familias conocidas. Dependiendo del número de
especies descritas para el Mundo (entre 110.000 y 150.000),
en el Neotrópico se ha descrito entre el 16 y el 22%
del total de especies conocidas.
Monografías y posibilidades de identificación
La
información sintetizada en el Anexo
I permite deducir la magnitud de monografías,
revisiones o sinopsis que facilitan la identificación
de himenópteros neotropicales hasta los niveles de
género o especie. En lo posible se citan referencias
recientes, desde la mitad de siglo. De las 77 familias relacionadas
en el Anexo, es posible la identificación a género
en 65, lo cual representa un 84%; en otras cinco (un 7%) se
puede llegar a género en, al menos, la mitad de las
subfamilias. Esto deja siete familias (9%) en donde no es
posible la identificación a niveles inferiores a subfamilia.
En 33 familias (43%) es posible, mediante literatura, la identificación
hasta nivel de especie, en otras 31 familias (40%), es posible
la identificación a especie de algunos grupos; el restante
número de familias no posee literatura apropiada para
este nivel de identificación.
El grupo mejor estudiado es el de las avispas-sierra (“Symphyta”),
pues las revisiones de D. Smith cubren todos los taxones neotropicales
hasta el nivel de especie. Actualmente Smith está revisando
la última y mayor familia, Tenthredinidae. Stephanoidea
está en revisión, mientras que en Trigonaloidea
sólo es posible llegar a género.
En la gigantesca superfamilia Ichneumonoidea es posible la
identificación de géneros en todas las subfamilias
de Braconidae y en algunas de Ichneumonidae.
La identificación hasta especie es posible en reducidos
grupos, pues la vasta mayoría de taxones en esta superfamilia
están sin estudiar: muchos ejemplares coleccionados
corresponden a especies sin describir.
En las superfamilias de Aculeata (Vespomorpha),
es factible la identificación a género en todas
las familias excepto Mutillidae, donde sólo existen
claves regionales para Panamá, Guyana y Argentina.
Por su tamaño (alrededor de 1500 especies), esta es
una familia donde es urgente un trabajo de revisión
al nivel de género. Aunque existen algunas monografías
parciales para Pompilidae, es otra familia donde se necesitan
claves integrales que cubran toda la región neotropical.
Numerosos géneros en esta división han sido
revisados (o están en revisión) especialmente
en hormigas, véspidos, abejas, "esfécidos"
y pompílidos.
Cynipoidea es la superfamilia más desconocida
de todo el orden. No sólo filogenéticamente
su posición es incierta, sino que se carece de consenso
en los límites de sus familias y subfamilias. El trabajo
de Weld (1952) está decididamente desfasado y las claves
para el mayor grupo en los trópicos (Eucoilidae
o Eucoilini) son muy difíciles de usar. Se necesitan
análisis filogenéticos en este grupo para definir
los géneros y proveer una clave moderna.
Proctotrupoidea es un grupo relativamente mejor conocido,
con tres familias pequeñas y dos cosmopolitas presentes
en la región. En una de éstas, Proctotrupidae,
existe una monografía que permite la identificación
de especies. En la otra, Diapriidae, Masner y García
están concluyendo la revisión genérica
de la subfamilia Diapriinae, la más común en
América Tropical. La mayoría de los géneros
están sin revisar.
En Platygastroidea es posible la identificación
hasta género en Scelionidae y gran parte de
Platygastridae, aunque en el primer caso pueden existir
varios géneros aún no descritos. Los géneros
neotropicales revisados son escasos.
Mymarommatidae, aunque pequeño (un género
viviente), es un grupo enigmático con apenas uno o
dos hallazgos en la región Neotropical.
Chalcidoidea presenta el gran problema de la definición
interna de sus familias, y es un taxón con grupos bien
estudiados (p.e. Chalcididae, Leucospidae o
Eupelmidae) a pobremente definidos y revisados (p.e.
Pteromalidae y Aphelinidae). Aunque es posible
llegar hasta el nivel de género en varios grupos importantes,
el tamaño de muchas de estas avispas y la escasez de
monografías hacen difíciles las identificaciones
de especies.
Megalyroidea es un grupo muy pequeño, con pocas
especies neotropicales revisadas recientemente.
Ceraphronoidea es otra de las superfamilias pobremente
estudiadas. Aunque es posible la identificación de
géneros, la taxonomía de las especies neotropicales
es un campo totalmente virgen.
Evanioidea es un grupo pobremente estudiado filogenética
y taxonómicamente. Es factible la identificación
genérica (por ser pocos los conocidos), pero, con la
excepción de Aulacidae (que está en revisión),
la identificación específica es prácticamente
imposible.
Catálogos
No
existen catálogos modernos para la fauna neotropical
de himenópteros y la información que se puede
obtener de los viejos catálogos mundiales es obsoleta.
Pagliano y Scaramozzino (1990) presentan un listado sinonímico
de los 17.000 géneros y subgéneros de Hymenoptera
descritos hasta esa fecha, incluyendo fósiles. Abe
y Smith (1990) presentan un catálogo para todas las
categorías supraespecíficas de “Symphyta”
del Mundo y Weinstein y Austin (1991) publicaron otro catálogo
abreviado para las especies de las Trigonalidae del
Mundo; Townes y Townes (1966) y Yu y Horstmann (1997a,b) presentan,
respectivamente, catálogos para las Ichneumonidae
del Neotrópico y el mundo. Gordh y Moczar (1990) ofrecen
el catálogo de las Bethylidae del mundo y en Kimsey
y Bohart (1990) se puede extraer el listado de especies de
las Chrysididae del mundo.
El tratado de Sphecidae de Bohart y Menke (1976) incluye
listados de especies para todo el mundo, así como el
listado sinonímico de todas las categorías por
encima de especie. Moure y Hurd (1987) ofrecen el catálogo
de las Halictidae del Hemisferio Occidental. Existe
el catálogo abreviado para las hormigas neotropicales
(Kempf, 1972; Brandao, 1991) y el catálogo para las
especies del Mundo (Bolton, 1995). Johnson (1992) y Vlug (1995)
ofrecen el catálogo de los Proctotrupoidea y
Platygastroidea del Mundo.
De Santis (1979, 1983, 1989) y De Santis y Fidalgo (1994 y
en preparación) publicaron el catálogo (y suplementos)
de los Chalcidoidea de América al sur de los
EE.UU. De Santis (1980) publicó también un catálogo
para los “Parasitica y Chrysidoidea”
de Brasil.
Superfamilias
y familias altamente diversas
Las
superfamilias más diversas en la región (con
más de 1000 especies) son Tenthredinoidea, Chrysidoidea,
Chalcidoidea, Ichneumonoidea, Apoidea y
Vespoidea, las tres últimas con más de
4.000 especies cada una (Tabla
II, Fig. 3).
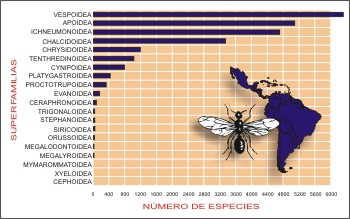
Fig.
3. Número de especies por superfamilia en
la región Neotropical
Las familias más diversas (en número de especies
descritas, tomando un número arbitrario mínimo
de 300) son Tenthredinidae, Bethylidae, Chrysididae,
Dryinidae, Colletidae, Andrenidae, Halictidae,
Megachilidae, Pompilidae y Encyrtidae. Familias muy
diversas, con más de 1.000 especies descritas para
la región, son Crabronidae, Apidae, Vespidae
y Mutillidae. Familias hiperdiversas, con más
de 2.000 especies en la región, son Braconidae,
Ichneumonidae y Formicidae.
Superfamilias diversas desde el punto de vista de número
de géneros, y asumiendo el número arbitrario
de 100, son Tenthredinoidea (115), Apoidea (352) y
Vespoidea (373) (Tabla II y Fig.
4). Superfamilias altamente diversas genéricamente
son Ichneumonoidea (694) y Chalcidoidea (625),
ambas con alrededor del 48% de todos los géneros neotropicales
de Hymenoptera.
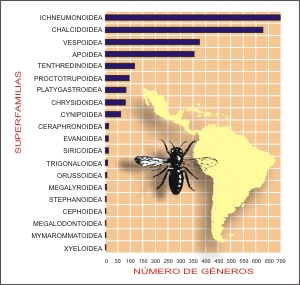
Fig.
4. Número de géneros por superfamilia
en la región Neotropical
Taxones
pobremente estudiados
Los
grupos peor conocidos en la región Neotropical son
Ceraphronoidea, Cynipoidea y Evanioidea.
Dentro de algunas estimaciones de números de especies,
Ichneumonoidea y demás grupos de parasitoides
están muy pobremente estudiados.
Especialistas
y Museos
Aparte
de los grandes museos que albergan muchos tipos y material
asociado coleccionado en los países tropicales (Museo
Nacional de Historia Natural de los EE.UU., Museo Americano
de Historia Natural en Nueva York, Museo de Historia Natural
de Londres, Colección Nacional del Canadá, etc.),
son pocos los museos iberoamericanos que guardan colecciones
importantes que representen la región Neotropical.
Incluso en varios países no existen museos que almacenen
porciones representativas de sus propias himenopterofaunas.
La colección de insectos del Museo de Zoología
en São Paulo está bien representada en varios
grupos de himenópteros (especialmente en hormigas).
El INBio de Costa Rica posee una gran colección de
himenópteros del país. En México, Brasil
y Argentina hay otras colecciones importantes. Nuevas colecciones
de varios grupos del orden se están fortaleciendo en
otros países como Venezuela y Colombia.
No existe un “inventario” de taxónomos
que total o parcialmente estén involucrados en el estudio
de la fauna neotropical. En grupos conspicuos como Formicidae
hay una veintena de profesionales y estudiantes que están
comprometidos en estudios de filogenia y revisiones de varias
tribus y géneros. Cifras semejantes pueden manejarse
para las abejas, los "esfécidos" y los calcidoideos.
Varias superfamilias tienen especialistas “solitarios”
(como las de “Symphyta”, Stephanoidea,
Trigonaloidea, Megalyroidea). Algunas otras
son inmensas y los pocos especialistas existentes son insuficientes
(como Ichneumonoidea); otras (como Ceraphronoidea)
están virtualmente huérfanas.
Perspectivas
El
siglo que se avecina trae consigo retos difíciles y
un panorama complicado. Hymenoptera ocupa, al parecer,
un segundo lugar entre los órdenes hiperdiversos de
insectos, con algo más de 250.000 especies estimadas
(Hanson y Gauld, 1995). La mayoría de estas especies
permanecen sin describir, y además una inmensa porción
de estos insectos vive en los trópicos. Probablemente
la región Neotropical albergue no menos de 60.000 especies,
con sólo una fracción descrita.
El estudio y comprensión de la himenopterofauna neotropical
se enfrenta a dos problemas: la escasez de taxónomos
y la disminución de los ecosistemas y hábitats
donde prospera la mayoría de especies. Varios especialistas
coinciden en que una fracción importante de esta fauna,
podría desaparecer antes aún de que pudieran
nombrarse.
Pero, si el solo hecho de colocar nombres es una tarea formidable
y prácticamente imposible, existen otras tareas igualmente
inmensas: estudiar y proponer las relaciones filogenéticas
de los grupos (que obviamente requiere de la comprensión
de la fauna mundial), entender sus patrones biogeográficos
y dilucidar su historia natural (Mound, 1998). Los últimos
factores son críticos para una mejor comprensión
de la biología de estos insectos en los trópicos
y, eventualmente, para su uso en estrategias de conservación,
control biológico, polinización, etc.
Mound y Gaston (1993) han sugerido que la taxonomía
descriptiva debe dejar de ser prioritaria en las agendas sistemáticas
de instituciones y universidades. Estos autores señalan
que aun con un alto número de taxónomos y de
recursos apropiados, se tardarían muchas décadas
tan solo nombrando taxones, describiéndolos y publicándolos.
Un problema paralelo está en la existencia de sitios
donde se puedan almacenar eficazmente los ejemplares y toda
la información asociada a éstos (Gauld, 1996).
No existen instituciones que puedan aceptar guardar tipos
y ejemplares de la gigantesca himenopterofauna tropical. Instituciones
grandes como los museos de Washington y Londres poseen pocos
taxónomos en Hymenoptera y algunos tratan parcialmente
la fauna neotropical o están dedicados a faunas de
otras regiones. El Museo de Londres está avocado a
serios recortes presupuestarios, lo cual ha obligado a recortes
de personal y/o la búsqueda de fuentes alternativas
de financiación.
Las instituciones latinoamericanas poseen problemas aún
más graves, ante la carencia de recursos y la aguda
escasez de investigadores en sistemática (Barros de
Carvalho, 1998). Es una gran paradoja, pues estos países
son los “depositarios naturales” de la megadiversidad
de artrópodos. Varias instituciones (especialmente
en Europa) no prestan los tipos correspondientes a la fauna
neotropical, lo cual encarece los costos de una revisión
o simplemente impiden su ejecución. En los centros
de enseñanza superior de una u otra forma se alimenta
la idea de que la ecología o la biología molecular
son más “interesantes y lucrativas” que
la sistemática, logrando, en algunos casos, desanimar
a jóvenes potencialmente interesados en taxonomía
(al fin y al cabo un taxónomo jamás recibirá
el Premio Nobel).
Un mundo superpoblado de ecólogos y biólogos
moleculares trabajando con organismos sin nombre es previsible
para las próximas generaciones. Por otra parte, es
interesante la reciente observación de que estas áreas
tan excitantes (como la biología molecular) no necesariamente
pueden estar generando nuevas ideas y paradigmas en la biología,
como la evolución y la sistemática en los siglos
XIX y XX: “una intensa concentración de recursos
en biología molecular podría estar desacelerando
el progreso conceptual en biología” (Vogel, 1998).
Quizá la taxonomía no sea una disciplina biológica
tan atractiva como otras, pero sigue estando en el problema
central del entendimiento y definición de la diversidad
biológica.
Agradecimiento
La
elaboración del presente diagnóstico ha sido
posible gracias a numerosos colegas y amigos que han ofrecido
ayuda en diversas formas. Entre ellos destaco a Paul Hanson
(Universidad de Costa Rica), Arnold S. Menke (Smithsonian
Institution), Lubomir Masner (Colección Nacional del
Canadá), Charles D. Michener (Universidad de Kansas),
José L. Nieves-Aldrey (Museo de Historia Natural, Madrid),
Massimo Olmi (Universidad de Tuscia, Italia), Luis de Santis
(Museo de La Plata), Michael Sharkey (Universidad de Kentucky)
y David Smith (Smithsonian Institution).
También agradezco a G. Halffter, F.Martín-Piera
y F. Escobar por su invitación a participar en el Proyecto,
y a Claudia Martínez, Carlos Sarmiento, Edgard Palacio,
Diego Campos, Juan Manuel Vargas y Mónica Ospina por
su ayuda y asesoría en sus grupos de trabajo y/o elaboración
de gráficas. Fermín Martín-Piera y un
revisor anónimo hicieron importantes comentarios al
manuscrito.
Bibliografía
ABE,
M. Y D. R. SMITH. 1991. The genus-group names of Symphyta
(Hymenoptera) and their type species. Esakia, 31: 1-115
ACHTERBERG, C. Y D. L. J. QUICKE. 1992. Phylogeny of the subfamilies
of the family Braconidae: A reassessment assessed. Cladistics,
8: 237-264
AGUIAR, A. P. 1998. Revisao do genero Hemistephanus Enderlein,
1906 (Hymenoptera, Stephanidae), com consideraçoes
metodológicas. Rev. Bras. Entomol., 41(2-4): 343-429
ALEXANDER, R. 1992. An exploratory analysis of cladistic relationships
within the superfamily Apoidea, with special reference to
sphecids wasps. J. Hym. Res., 1: 25-61
ALEXANDER, R. Y C. D. MICHENER. 1995. Phylogenetic studies
of the families of short-tongued bees. Univ. Kansas Sci. Bull.,
55: 377-424
ALLEN, H. W. 1972. A monographic study of the subfamily Tiphiinae
(Hymenoptera: Tiphidae) of South America. Smithson. Contrib.
Zool., 113: 1-76
AMARANTE, S. T. P., C. R. F. BRANDAO Y J. M. CARPENTER. 1999.
A new species of Embolemus Westwood from the Mata Atlantica
of Brazil (Hymenoptera: Embolemidae). Am. Mus. Novit., 3266:
1-7
ARGAMAN, Q. 1988. Generic synopsis of Sclerogibbidae (Hymenoptera).
Ann. Hist.-Nat. Mus. Nat. Hung., 80: 177-187
AZEVEDO, C. O. 1999. A key to world species of Scolebythydae
(Hymenoptera: Chrysidoidea), with description of a new species
of Dominibythus from Brasil. J. Hym. Res., 8(1): 1-5
BANKS, N. 1946. Studies of South American Psammocharidae.
Part I. Bull. Mus. Comp. Zool., 96: 311-525
BANKS, N. 1947. Studies of South American Psammocharidae.
Part II. Bull. Mus. Comp. Zool., 99: 371-486
BARONI, C., B. BOLTON Y P.S. WARD. 1992. The internal phylogeny
of ants (Hymenoptera: Formicidae). Syst. Entomol., 17: 301-329
BARROS DE CARVALHO, C. J. 1998. Taxonomista de insetos, una
especie em extinçao no Brasil. Info. Soc. Entomol.
Brasil, 23(1): 1,5
BASIBUYUK, H.H. Y D.L.J. QUICKE. 1995. Morphology of the antenna
cleaner in the Hymenoptera with particular reference to non-aculeate
families (Insecta). Zool. Scripta, 24 (1-2): 157-177
BASIBUYUK, H.H. Y D.L.J. QUICKE. 1999. Gross morphology of
multiporous plate sensilla in the Hymenoptera (Insecta). Zool.
Scripta, 28(1-2): 51-67
BELSHAW, R., M. FITTON, E. HERNIOU, C. GIMENO Y D.L.J. QUICKE.
1998. A phylogenetic reconstruction of the Ichneumonoidea
(Hymenoptera) based on the D2 variable region of 28S ribosomal
RNA. Syst. Entomol., 23: 109-123
BOHART, R. M. Y A.S. MENKE. 1976. Sphecid wasps of the World,
a generic revision. University of California Press, Berkeley.
BOLTON, B. 1994. Identification Guide to the Ant Genera of
the World. Harvard University Press, Cambridge.
BOLTON, B. 1995. A New general Catalogue of the Ants of the
World. Harvard University Press, Cambridge.
BORROR, D.J., C.A. TRIPLEHORN Y N.F. JOHNSON. 1989. An introduction
to the study of insects. Saunders College Publishing.
BOUCEK, Z. 1974. A revision of the Leucospidae (Hymenoptera:
Chalcidoidea) of the World. Bull. Brit. Mus. (Nat. Hist.),
Entomol. Suppl., 23: 1-241
BOUCEK, Z. 1988. An overview of the higher classification
of the Chalcidoidea (Parasitic Hymenoptera). In: GUPTA, V.
K. (ed.), Advances in Parasitic Hymenoptera Research, E.J.
Brill, Leiden. pp. 11-23
BOUCEK, Z. 1992. The New World genera of Chalcididae. In:
DELVARE, G. Y BOUCEK, Z. On the New World Chalcididae (Hymenoptera).
Mem. Amer. Entomol. Inst., 53, pp. 49-117.
BOUCEK, Z. Y J. S. NOYES. 1987. Rotoitidae, a curious new
family of Chalcidoidea (Hymenoptera) from New Zealand. Syst.
Entomol., 12: 407-412
BRADLEY, J. C. 1945. The Scoliidae (Hymenoptera) of norhern
South America, with especial reference to Venezuela. I. The
genus Campsomeris. Bol. Entomol. Venez., 4(1): 1-36
BRANDAO, C. R. F. 1991 Adendos ao catálogo abreviado
das formigas da regiao Neotropical. Revta. Bras. Entomol.,
35: 319-412
BRANDAO, C. R. F., J. L. M. DINIZ, D. AGOSTI Y J. H. DELABIE.
1999. Revision of the Neotropical Ant Subfamily Leptanilloidinae.
Syst. Entomol., 24(1): 17-36
BROTHERS, D. J. 1975. Phylogeny and Classification of the
Aculate Hymenoptera, with special reference to Mutillidae.
Univ. Kan. Sci. Bull., 50(11): 483-648
BROTHERS, D. J. 1999. Phylogeny and Evolution of wasps, ants
and bees (Hymenoptera, Chrysidoidea, Vespoidea and Apoidea).
Zool. Scripta, 28(1-2): 233-249
BROTHERS, D. J. Y J. M. CARPENTER. 1993. Phylogeny of Aculeata:
Chrysidoidea and Vespoidea (Hymenoptera). J. Hym. Res., 2:
227-302
BROWN, K.S. 1991 Conservation of Neotropical Environments:
Insects as indicators. In: COLLINS, N. M. Y J. A. THOMAS (eds.),
Conservation of Insects and their Environments Academic Press,
Londres, pp. 349-404.
CARMEAN, D. Y L. S. KIMSEY. 1998. Phylogenetic revision of
the parasitoid wasp family Trigonalidae (Hymenoptera). Syst.
Entomol., 23: 35-76
CARPENTER, J. M. 1982. The phylogenetic relationships and
natural classification of the Vespoidea (Hymenoptera). Syst.
Entomol., 7: 11-38
CARPENTER, J. M. 1986a. Cladistics of the Chrysidoidea (Hymenoptera).
J. New York Entomol. Soc., 94: 303-330
CARPENTER, J. M. 1986b. A Synonimic generic cheklist of the
Eumeninae (Hymenoptera: Vespidae). Psyche, 93: 61-90
CARPENTER, J. M. 1989 The phylogenetic system of the Gayellini
(Hymenoptera: Vespidae; Masarinae). Psyche, 95: 211-241
CARPENTER, J. M. 1999. What do we known about chrysidoid relationships?
Zool. Scripta, 28(1-2): 215-231
CARPENTER, J. M. Y W. C. WHEELER. 1999 Toward simultaneous
analysis of morphological and molecular data in Hymenoptera.
Zool. Scripta, 28(1-2): 251-260
CROZIER, R. H. 1977 Evolutionary genetics of the Hymenoptera.
Annu. Rev. Entomol., 22: 263-288
DELVARE, G y Boucek, Z. 1992. On the New World Chalcididae
(Hymenoptera). Mem. Amer. Entomol. Inst., 53: 1-466
DE SANTIS, L. 1979. Catálogo de los Himenópteros
Calcidoideos de América al sur de los Estados Unidos.
Pub. Esp. Com. Invest. CiEntomol. Prov. Buenos Aires, La Plata.
DE SANTIS, L. 1983. Catálogo de los himenópteros
Calcidoideos de América al sur de los Estados Unidos
– Primer Suplemento. Rev. Peru. Entomol., 24: 1-38
DE SANTIS, L. 1989. Catálogo de los himenópteros
Calcidoideos de América al sur de los Estados Unidos
– Segundo Suplemento. Acta Entomol. Chil., 15: 9-90
DE SANTIS, L. Y P. FIDALGO. 1994. Catalogo de Himenópteros
Calcidoideos. Serie No. 13, Academia Nacional de Agronomía
y Veterinaria, Buenos Aires.
DESSART, P. Y P. CANSEMI. 1986. Tableau dichotomique des genres
de Ceraphronoidea (Hymenoptera) avec commentaires et nouvelles
espèces. Frust. Entomol. (N.S), 7-8: 307-372.
DOWTON, M. Y A. D. AUSTIN. 1994. Molecular phylogeny of the
insect order Hymenoptera: Apocritan relationships. Proc. Natl.
Acad. Sci. USA, 91: 9911-9915.
DOWTON, M. Y A. D. AUSTIN. 1999. Models of analysis for Molecular
data sets for the Reconstruction of Basal Hymenopteran Relationships.
Zool. Scripta, 28(1-2): 69-74.
DOWTON, M., A. D. AUSTIN, N. DILLON Y E. BARTOWSKY. 1997.
Molecular phylogeny of the apocritan wasps: the Proctotrupomorpha
and Evaniomorpha. Syst. Entomol., 22: 245-255.
EADY, R. D. 1968. Some Illustrations of Microsculpture in
the Hymenoptera. Proc. Royal Entomol. Soc. London (A), 43(3-6):
66-72.
ELLIOT, E. A. 1922. Monograph of the hymenopterous family
Stephanidae. Proc. Zool. Soc. London, 1922: 705-831.
EVANS, H. E. 1961. A preliminary review of the Nearctic species
of Sierolomorpha (Hymenoptera). Breviora, 140: 1-12.
EVANS, H. E. 1963. A new family of wasps. Psyche, 70: 7-16.
EVANS, H. E. 1964. A synopsis of the American Bethylidae (Hymenoptera:
Aculeata). Bull. Mus. Comp. Zool., 132: 1-122.
EVANS, H. E. 1966. A revision of the Mexican and Central American
spider wasps of the subfamily Pompilinae (Hymenoptera: Pompilidae).
Mem. Amer. Entomol. Soc., 20: 1-442.
EVANS, H. E. 1987. Order Hymenoptera. En: STEHR, F.W. (ed.),
Immature Insects, Vol. 1, Kendall/Hunt Pub. Co., Iowa, pp.
597-710.
FERGUSSON, N. D. M. 1995. The Cynipoid Family. En: HANSON,
P. Y I.D. GAULD (eds) en: Hymenoptera of Costa Rica. Oxford
University Press London, pp 247-265.
FERNÁNDEZ, F., M. RODRÍGUEZ Y G. ULLOA. 1996.
Primer registro de la Familia Sclerogibbidae para Colombia.
Sphecos, 30: 10.
GASTON, K. 1993. Spatial patterns in the description and richness
of the Hymenoptera. En LA SALLE, J. Y I.D. GAULD (eds) Hymenoptera
and Biodiversity, International Press, UK: pp. 277-293
GASTON, K., I.D. GAULD Y P. HANSON. 1996. The size and composition
of the hymenopteran fauna of Costa Rica. J. Biogeogr., 23:
105-113
GAULD, I. 1996. The cost of data repatriation versus the potential
of Biosystematic capacity building. En: MCNEELY, J.A. Y S.
SOUCHERITA (eds) Biodiversity in Asia: Challenges and Opportunities
for the Scientific Community. Ministry of Science, Technology
and Environment, Bangkok.
GAULD, I.D. Y B. BOLTON (eds.). 1988. The Hymenoptera. Oxford
University Press, Oxford.
GAULD, I.D. Y J. LA SALLE (eds.). 1993. Hymenoptera and Biodiversity.
C.A.B. International Press.
GENISE, J.F. 1984. Las Anthoboscinae neotropicales y comentarios
sobre la diversidad de la subfamilia (Hymenoptera, Tiphiidae).
Rev. Soc. Entomol. Arg., 43: 195-220.
GENISE, J.F. 1986. Las Bradynobaenidae y algunas modificaciones
a la clasificación general de Hymenoptera Aculeata.
Physis (Buenos Aires), C, 44: 39-53
GIBSON, G.A.P. 1985. Some pro and mesothoracic structures
important for phylogenetic analysis of Hymenoptera, with a
review of terms used for the structure. Can. Entomol., 117:
1395-1443.
GIBSON, G.A.P. 1986. Evidence for monophyly and relationships
of Chalcidoidea, Mymaridae, and Mymarommatidae (Hymenoptera:
Terebrantes). Can. Entomol., 118: 205-240.
GIBSON, G.A.P. 1989. Phylogeny and classification of Eupelmidae,
with a revision of the world genera of Calosotinae and Metapelmatinae
(Hymenoptera: Chalcidoidea). Mem. Entomol. Soc. Canada, nº
149. 121 pp.
GIBSON, G.A.P. 1993. Groundplan structure and homology of
the pleuron in Hymenoptera based on a comparison of the skeletomusculature
of Xyelidae (Hymenoptera) and Raphidiidae (Neuroptera). Mem.
Entomol. Soc. Can., 165: 165-187.
GIBSON, G.A.P. 1999. Sister-group relationships of the Platygastroidea
and Chalcidoidea (Hymenoptera) – an alternate hypothesis
to Rasnitsyn (1988). Zool. Scripta, 28(1-2): 125-138.
GIBSON, G.A.P., J.M. HERATY Y J.B. WOOLLEY. 1999. Phylogenetics
and classification of Chalcidoidea and Mymarommatoidea –
a review of current concepts (Hymenoptera, Apocrita). Zool.
Scripta, 28(1-2): 87-124.
GODFRAY, H.C.J. 1994. Parasitoids: Behavioral and Evolutionary
Ecology. Princeton University Press, NJ. Princeton University
Press.
GORDH, H. Y L. MOCZAR. 1990. A catalog of the world Bethylidae
(Hymenoptera: Aculeata). Mem. Amer. Entomol. Inst., 46: 1-364.
GOULET, H. Y J. HUBER (eds). 1993. Hymenoptera of the World:
An identification guide to families. Agriculture Canada, Ottawa.
GRIMALDI, D., D. AGOSTI Y J.M. CARPENTER. 1997. New and rediscovered
primitive ants (Hymenoptera: Formicidae) in cretaceous amber
from New Jersey, and their phylogenetic relationships. Am.
Mus. Nov., 3208: 1-43.
GUGLIELMINO, A. Y M. OLMI. 1997. A host-parasite catalog of
world Dryinidae (Hymenoptera: Chrysidoidea). Contr. Entomol,
2(2): 165-298.
HANSON, P. 1992. The nearctic species of Ormyrus Westwood
(Hymenoptera: Chalcidoidea: Ormyridae). J. Nat. Hist., 26:
1333-1365.
HANSON, P. Y I.D. GAULD (eds.). 1995 Hymenoptera of Costa
Rica. Oxford University Press, Oxford.
HARRIS, R.A. 1979. A Glossary of Surface Sculpturing. Occasional
Papers in Entomology (Davis) No. 28, 31 pp.
HAWKINS, B.A. Y G. GORDH. 1986. Bibliography of the World
Literature of the Bethylidae (Hymenoptera: Bethyloidea). Insecta
Mundi, 1(4): 261-283.
HERATY, J.M. Y D.C. DARLING. 1984. Comparative Morphology
of the planidial larvae of Eucharidae and Perilampidae (Hymenoptera:
Chalcidoidea). Syst. Entomol., 9: 309-328.
HERATY, J.M., J.B. WOOLLEY Y D.C. DARLING. 1994. Phylogenetic
implications of the mesofurca and mesopostnotum in Hymenoptera.
J. Hym. Res., 3: 241-277.
HÖLLDOBLER, B. Y E.O. WILSON. 1990. The Ants. Harvard
University Press.
HUBER, J.T. 1986. Systematics, biology, and hosts of the Mymaridae
and Mymarommatidae (Insecta: Hymenoptera): 1758-1984. Entomography,
4: 185-243.
ISIDORO, N., F. BIN, R. ROMANI, J. PUJADE Y P. ROS. 1999.
Diversity and function of male antennal glands in Cynipoidea
(Hymenoptera). Zool. Scripta, 28(1-2): 165-174.
JOHNSON, N.F. 1988. Midcoxal articulations and the phylogeny
of the order Hymenoptera. Ann. Entomol. Soc. Am., 81: 870-881
JOHNSON, N.F. 1992. Catalog of world species of Proctotrupoidea,
exclusive of Platygastridae (Hymenoptera). Mem. Amer. Entomol.
Inst., 51: 1-825.
KEMPF, W.W. 1972. Catálogo abreviado das formigas da
regiao neotropical (Hymemoptera: Formicidae). Stud. Entomol.,
15: 3-344.
KIMSEY, L.S. 1991. Relationships among the tiphiid wasp subfamilies
(Hymenoptera). Syst. Entomol., 16: 427-438
KIMSEY, L.S. 1992. Phylogenetic relations among the South
American thynnine tiphiid wasps (Hymenoptera). Syst. Entomol.,
17: 133-144.
KIMSEY, L.S. 1993. An unusual new tiphiid genus from Peru
and a key to American genera of Tiphiinae (Hymenoptera). Pan.
Pac. Entomol., 69: 213-217.
KIMSEY, L.S. Y R. M. BOHART. 1990. The Chrysididae Wasps of
the World. Oxford University Press, Oxford.
KÖNIGSMANN, E. 1978. Das phylogenetische System der Hymenoptera.
Teil 3. Terebrantes (Unterordnung Apocrita). Dtsch. Entomol.
Z., 25: 1-55.
KRISTENSEN, N.P. 1991. Phylogeny of Extant Hexapods. En: The
Insects of Australia CSIRO Pub, Camberra, pp. 125-140.
KROMBEIN, K.V. 1937. Studies in the Tiphiidae (Hymenoptera:
Aculeata). I. A review of the genera of Myzininae. An. Entomol.
Soc. Amer., 30: 26-30.
KROMBEIN, K.V. 1979. Studies in the Tiphiidae. XII. A new
genus of Methocinae with notes on the subgenera of Methoca
Latreille (Hymenoptera Aculeata). Proc. Entomol. Soc. Was.,
81: 424-434.
LA SALLE, J. 1987. New World Tanaostigmatidae (Hymenoptera,
Chalcidoidea). Cont. Amer. Entomol. Inst., 23(1): 1-181.
LA SALLE, J. Y I.D. GAULD. 1991. Parasitic Hymenoptera and
the Biodiversity Crisis. Redia, 76(3): 315-334.
LA SALLE, J. Y I.D. GAULD (eds). 1993. Hymenoptera and Biodiversity.
CAB International, UK.
MALAISE, R. 1963. Hymenoptera Tenthredinoidea, subfamiliy
Selandriinae, key to the genera of the world. Entomol. Z.,
84: 159-215.
MASNER, L. 1976a. A revision of the Ismarinae of the New World
(Hymenoptera: Proctotrupoidea, Diapriidae). Can. Entomol.,
108: 1243-1266.
MASNER, L. 1976b. Revisionary notes and keys to world genera
of Scelionidae (Hymenoptera: Proctotrupoidea). Mem. Entomol.
Soc. Can., 97: 1-87.
MASNER, L. 1980. Key to the Holarctic genera of Scelionidae,
with descriptions of new genera and species (Hymenoptera:
Proctotrupoidea). Mem. Entomol. Soc.Can., 113: 1-54.
MASNER, L. Y P. DESSART. 1967. La reclassification des categories
taxonomiques superieures des Ceraphronoidea (Hymenoptera).
Bull. Inst. R. Sci. Nat. Belg., 43: 1-33.
MASNER, L. Y L. HUGGERT. 1989. World review and keys to genera
of the subfamily Inostemmatinae with reassignment of the taxa
to the Platygastrinae and Sceliotrachelinae (Hymenoptera:
Platygastridae). Mem. Entomol. Soc. Can., 147: 1-214.
MELO, A.R. 1999. Phylogenetic relationships and classification
of then major lineages of Apoidea (Hymenoptera), with emphasis
on the crabronid wasp. Sci. Pap., Nat. Hist. Mus., Univ. of
Kansas, 14: 1-55.
MENKE, A.S. Y F. FERNÁNDEZ. 1996. Claves ilustradas
para las subfamilias, tribus y géneros de esfécidos
neotropicales (Apoidea: Sphecidae). Rev. Biol. Trop., Vol.
44, suppl. 2, 1-68.
MICHENER, C.D. 1997. Genus-Group Names of Bees and Supplemental
Family-Group Names. Nat. Hist. Mus. Univ. Kan. Sci. Entomol.
Pap., 1: 1-81.
MICHENER, C.D. 2000. Bees of the World. John Hopkins University
Press.
MICHENER, C.D., R.J. MCGINLEY Y B.N. DANFORTH. 1994. The Bee
Genera of North and Central America (Hymenoptera: Apoidea).
Smithsonian Institution Press, Washington D.C.
MINELLI, A. 1993. Biological Systematics: The State of the
Art Chapman y Hall, London.
MOUND, L.A. 1998. Insect Taxonomy in Species-Rich Countries
- The Way Forward? An. Soc. Entomol. Bras., 27(1): 1-8
MOUND, L.A. Y K.J. GASTON. 1993. Conservation and systematics
– the agony and the ecstasy. En: GASTON, K.J., T.R.
NEW Y M.J. SAMWAYS (eds). Perspectives on Insect Conservation.
Intercept, UK, pp. 185-195
MOURE, J.S. Y HURD P.D., JR. 1987. An Annotated Catalog of
the Halictid Bees of the Western Hemisphere (Hymenoptera:
Halictidae). Smithsonian Institution Press Whashington D.C.,
405 pp.
NAUMANN, I.D. Y L. MASNER. 1985. Parasitic wasps of the proctotrupoid
complex: a new family from Australia and a key to world families
(Hymenoptera: Proctotrupoidea sensu lato). Aust. J. Zool.,
33: 761-783
NIEVES-ALDREY, J.L. Y F.M. FONTAL-CAZALLA. 1999. Filogenia
y Evolución del Orden Hymenoptera Bol. SEA, 26: 459-474
NONVELLIER, G. 1990. Catalogue of the Mutillidae, Myrmosidae
and Bradynobaenidae of the Neotropical Region including Mexico
(Insecta: Hymenoptera). Hymenopterum Catalogus Pars 18, SPB
Academic Publishing, Netherlands.
NORLANDER, G., Z. LIU Y F. RONQUIST. 1996. Phylogeny and historical
biogeography of the cynipoid wasp family Ibaliidae (Hymenoptera).
Syst. Entomol., 21: 151-166.
NOYES, J.S. 1980. A review of the genera of Neotropical Encyrtidae
(Hymenoptera: Chalcidoidea). Bull. Brit. Mus., 4(3): 107-253.
OLMI, M. 1984. A revision of the Drynidae (Hymenoptera). Mem.
Amer. Entomol. Inst., 37: 1-1913.
OLMI, M. 1986. New Species of Drynidae (Hymenoptera, Chrysidoidea).
Boll. Mus. Reg. Sci. Nat. Tor., 5: 211-238.
OLMI, M. 1989. Supplement to the revision of the world Drynidae
(Hymenoptera Chrysidoidea). Frust. Entomol. (N.S.),12: 109-395.
OLMI, M. 1993a. Drynidae di Costa Rica: catalogo e consideazioni
biogeografiche de evollutive (Hymenoptera Chrysidoidea). Boll.
Soc. Entomol. Ital., 124: 186-200.
OLMI, M. 1993b. A new generic classification for Thaumatodryininae,
Dryininae and Gonatopodinae, with descriptions of new species
(Hymenoptera Drynidae). Boll. Zool. Agr. Bachic., (Ser. II)
25: 57-89.
OLMI, M. 1995a. Contribution to the knowledge of the world
Dryinidae (Hymenoptera Chrysidoidea). Phytophaga, 6: 3-54.
OLMI, M. 1995b. A revision of the World Embolemidae (Hymenoptera
Chrysidoidea). Frust. Entomol., 8(31): 85-146.
PAGLIANO, G. Y P. SCARAMOZZINO. 1990. Elenco del Generi di
Hymenoptera del Mondo. Mem. Soc. Entomol. Ital., 68: 1-210.
PATE, V.S.L. 1947. Neotropical Sapygidae with a conspectus
of the family (Hymenoptera Aculeata). Acta Zool. Lilloana,
4: 393-426.
PERFECTO, I., J. VANDERMER, P. HANSON Y V. CARTÍN.
1997. Arthropod biodiversity loss and the transformation of
a tropical agro-ecosystems. Biodiver. Conserv., 6: 935-945.
QUICKE, D. L. J. 1997. Parasitic Wasps. Chapman & Hall,
London.
QUICKE, D.L.J. 1998. Parasitic Wasps Blackwell, London.
QUICKE, D.L.J., M.G. FITTON Y S. INGRAM. 1992a. Phylogenetic
implications of the structure and distribution of ovipositor
valvilli in the Hymenoptera (Insecta). J. Nat. Hist., 26:
587-608.
QUICKE, D.L.J., S.N. INGRAM, H.S. BAILLIE Y P.V. GAITENS.
1992b. Sperm structure and ultraestructure in the Hymenoptera
(Insecta). Zool. Scr., 21: 381-402.
QUICKE, D.L.J., M.G. FITTON, J.R. TUNSTEAD, S.N. INGRAM Y
P.V. GAITENS. 1994. Ovipositor structure and relationships
within the Hymenoptera, with special reference to the Ichneumonoidea.
J. Nat.Hist., 28: 635-682.
QUICKE, D.L.J., H.H. BASIBUYUK, M.G. FITTON Y A.P. RASNITSYN.
1999a. Morphological, Paleontological and molecular aspects
of ichneumonoid phylogeny (Hymenoptera, Insecta). Zool. Scripta,
28(1-2): 175-202.
QUICKE, D.L.J., R. BELSHAW Y C. LÓPEZ-VAAMONDE. 1999b.
Preservation of hymenopteran specimens for subsequent molecular
and morphological study. Zool. Scripta, 28(1-2): 261-267.
QUICKE, D.L.J., C. LÓPEZ-VAAMONDE Y R. BELSHAW. 1999c.
The Basal Ichneumonidae (Insecta, Hymenoptera): 28S D2 rDNA
considerations of the Brachycyrtinae, Labeninae, Paxylommatinae
and Xoridinae. Zool. Scripta, 28(1-2): 203-210.
QUINTERO, D. Y R. RODRIGUEZ. 1990. New Species of Typhoctes
from Panama (Hymenoptera: Typhoctidae). Ann. Entomol. Soc.
Am., 83(1): 30-35.
RAMIREZ, B.W. 1970. Taxonomic an biological studies of Neotropical
fig wasps (Hymenoptera: Agaonidae). Univ. Kan. Sci. Bull.,
49: 1-44.
RASNITSYN, A.P. 1988. An outline of the evolution of the hymenopterous
insects (Order Vespida). Oriental Insects, 22: 115-145.
RICHARDS, O.W. 1939. The Bethylidae subfamily Sclerogibidae
(Hymenoptera). Proc. R. Entomol. Soc. London, Series B 8:
211-223.
RICHARDS, O.W. 1962. A revisional study of the masarid wasps
(Hymenoptera: Vespidae) British Museum (Natural History),
London, England, 302p.
RICHARDS, O.W. 1978. The Social wasps of the Americas exluding
the Vespinae. British Museum (Natural History), London, 580
pp.
ROIG-ALSINA, A. 1994. A new genus of Plumariidae, with notes
on the relationships among the genera of the family (Hymenoptera,
Chrysidoidea, Plumariidae). Mitt. Münch. Entomol. Ges.,
84: 91-96.
ROIG-ALSINA, A. Y C.D. MICHENER. 1993. Studies of the phylogeny
and classification of long-tongued bees. Univ. Kan. Sci. Bull.,
55: 124-162.
RONQUIST, F. 1995b. Phylogeny and classification of the Liopteridae,
an archaic group of cynipoid wasps (Hymenoptera). Entomol.
Scand., 46: 1-74.
RONQUIST, F. 1995. Phylogeny and the early evolution of the
Cynipoidea (Hymenoptera). Syst. Entomol., 20: 309-335.
RONQUIST, F. 1999a. Phylogeny of the Hymenoptera: The state
of the art. Zool. Scripta, 28(1-2): 3-11.
RONQUIST, F. 1999b. Phylogeny, classification, and evolution
of the Cynipoidea. Zool. Scripta, 28(1-2): 139-164.
RONQUIST, A., A.P. RASNITSYN, A. ROY, K. ERIKSON Y M. LINDGREN.
1999. Phylogeny of the Hymenoptera: A cladistic reanalysis
of Rasnitsyn’s (1988) data. Zool. Scripta, 28(1-2):
13-50.
ROUBIK, D. 1989. Ecology and Natural History of Tropical Bees
Cambridge University Press.
SARMIENTO, C. 1999. Distribución de las Avispas Sociales
en Colombia (Hymenoptera: Vespidae: Polistinae) Biogeographica,
75(1): 17-36.
SHARKEY, M.J. y D.B. WAHL. 1992. Cladistics of the Ichneumonoidea
(Hymenoptera). J. Hym. Res.,1: 15-24.
SHATTUCK, S.O. 1992. Generic revision of the ant subfamily
Dolichoderinae. Sociobiology, 21: 1-181.
SHATTUCK, S.O. 1994. Taxonomic catalog of the ant subfamilies
Aneuretinae and Dolichoderinae. Univ. Cal. Pub. Entomol.,
112: 1-241.
SHAW, S.R. 1988. Euphorinae phylogeny: the evolution of diversity
in host-utilization by parasitoid wasps (Hymenoptera Braconidae).
Ecol. Entomol., 13: 323-335.
SHAW, S.R. 1990. Phylogeny and Biogeography of the parasitoid
wasp family Megalyridae (Hymenoptera). J. Biogeogr., 17: 569-581.
SHIMIZU, A. 1994. Phylogeny and Classification of the family
Pompilidae (Hymenoptera). Bull. Nat. Hist., Tokyo Met. Univ.,
2: 1-142.
SMITH, D.R. 1973. Sawflies of the subfamily Heterarthrinae:
adults and larvae (Hymenoptera: Tenthredinidae). Proc. Entomol.
Soc. Wash., 75: 337-345.
SMITH, D.R. 1978. Suborder Symphyta (Xyelidae, Pararchexyelidae,
Parapamphiliidae, Xyelydidae, Karatavitidae, Gigasiricidae,
Sepulcidae, Pseudosiricidae, Anaxyelidae, Siricidae, Xiphydriidae,
Paroryssidae, Xyelotomidae, Blasticotomidae, Pergidae). Hymenopterorum
Catalogus, Pars 14. 193 pp.
SMITH, D.R. 1988. A synopsis of the sawflies (Hymenoptera:
Symphyta) of America south of the United States: introduction,
Xyelidae, Pamphilidae, Cimbicidae, Diprionidae, Xiphydriidae,
Siricidae, Orussidae, Cephidae. Syst. Entomol., 13: 205-261.
SMITH, D.R. 1990. A synopsis of the sawflies (Hymenoptera,
Symphyta) of America south of the United States: Pergidae.
Rev. Bras. Entomol., 34(1): 7-200.
SMITH, D.R. 1992. A synopsis of the sawflies (Hymenoptera:
Symphyta) of America south of the United Sates: Argidae. Mem.
Entomol. Soc., 39: 1-119.
SMITH, D.R. 1993. Systematics, life history, and distribution
of sawflies. En: WAGNER, M. R. Y K.F. RAFFA (eds) Sawfly Life
History Adaptations to Woody Plants, Academic Press, pp. 3-32
.
STOLZ, D. Y J.B. WHITFIELD. 1992. Viruses and virus like entities
in the parasitic Hymenoptera. J. Hym. Res., 1: 125-139.
TONAPI, G.T. 1958a. A comparative study of spiracular structure
and mechanisms in some Hymenoptera. Trans. R. Entomol. Soc.,
110: 489-532.
TONAPI, G.T. 1958b. A comparative study of spiracular structure
and mechanisms in some Hymenoptera Part I: Symphyta. Indian
J. Entomol., 20: 108-120.
TONAPI, G.T. 1958c. A comparative study of spiracular structure
and mechanisms in some Hymenoptera Part II: Apocrita Parasitica.
Indian J. Entomol., 20: 203-220.
TONAPI, G.T. 1958d . A comparative study of spiracular structure
and mechanisms in some Hymenoptera Part III: Apocrita Aculeata.
Indian J. Entomol., 20: 245-269.
TOWNES, H. 1969. Genera of Ichneumonidae. Part 1. Mem. Amer.
Entomol. Inst., 11: 1-300.
TOWNES, H. 1970a.Genera of Ichneumonidae. Part 2. Mem. Amer.
Entomol. Inst., 12: 1-537.
TOWNES, H. 1970b. Genera of Ichneumonidae. Part 3. Mem. Amer.
Entomol. Inst., 13: 1-307.
TOWNES, H. 1971. Genera of Ichneumonidae. 4. Memoirs of The
American Entomological Institute, 17: 1-372.
TOWNES, H 1977a. A Revision of the Rhopalosomatidae (Hymenoptera).
Cont. Amer. Entomol. Inst., 15(1): 1-34.
TOWNES, H 1977b. A Revision of the Heloridae (Hymenoptera).
Cont. Amer. Entomol. Inst., 15(2): 1-12.
TOWNES, H. Y M. TOWNES. 1966. A catalogue and reclassification
of Neotropic Ichneumonidae. Mem. Amer. Entomol. Ins., 8: 1-367.
TOWNES, H. y M. TOWNES. 1981. A revision of the Serphidae
(Hymenoptera). Mem. Amer. Entomol. Inst., 32: 1-541.
VIHELMSEN, L. 1997a. Head capsule concavities accommodating
the antennal bases in Hymenoptera pupating in wood: possible
emergence-facilitating adaptations. Int. J. Ins. Morph. Embryol.,
26: 129-138
VIHELMSEN, L. 1997b. The phylogeny of lower Hymenoptera (Insecta),
with a summary of the early evolutionary history of the order.
J. Zool. Syst. Evol. Res., 35: 49-70.
VIHELMSEN, L.B. 1999. Comparative anatomy in the basal Hymenoptera
(Insecta), with a reappraisal of the occipital region. Zool.
Scripta, 28(1-2):75-85
VLUG, H.J. 1995. Catalogue of the Platygastridae (Platygastroidea)
of the World. En: C. VAN ACHTERBERG (DE) Hymenopterorum Catalogus
(Nova Editidio) Pars 19. Amsterdam, SBP Academic Publishing,
168 pp.
VOGEL, S. 1998. Academically Correct Biological Science. Amer.
Sci., 86(6): 504-506.
WARD, P.S. 1990. The ant subfamily Pseudomyrmecinae (Hymenoptera:
Formicidae) generic revision and relationship to other formicids.
Syst. Entomol., 15: 449-489.
WASBAUER, M.S. 1968. New genera of male Brachycistidinae with
redescription of Brachycistellus Baker and a key to North
American genera. Pan-Pacific Entomol., 44: 184-197.
WATKINS, J.F. 1976. The identification and Distribution of
New World Army Ants (Dorylinae: Formicidae), Baylor University
Press Waco Texas, 102 pp.
WELD, L.H., 1952. Cynipoidea (Hymenoptera) 1905 – 1950.
Privately printed Ann. Arbor, Michigan, 351 pp.
WEINSTEIN, P. Y A.D. AUSTIN. 1991. The host relationships
of trigonalyid wasps (Hymenoptera: Trigonalyidae), with a
review of their biology and catalogue to world species. J.
Nat. Hist., 25: 399-433.
WHARTON, R.A., P.M. MARSH Y M.J. SHARKEY. (eds) 1997 Manual
of the New World genera of the Family Braconidae (Hymenoptera).
Spec. Pub. No. 1, The International Society of Hymenopterists,
Washington D.C.
WHITFIELD, J.B. 1992. Phylogeny of the non-aculeate Apocrita
and the evolution of parasitism in the Hymenoptera. J. Hym.
Res., 1: 3-14.
WHITFIELD, J.B. 1998. Phylogeny and evolution of Host-Parasitoid
interactions in Hymenoptera. Ann. Rev. Entomol., 43: 129-151.
WHITFIELD, J.B., N.F. JOHNSON, Y M.R. HAMERSKI. 1989. Identity
and phylogenetic significance of the Metapostnotum in non-aculeate
Hymenoptera. Ann. Entomol. Soc. Am., 82: 663-674.
WILSON, E.O. 1971. The Insect Societes. Harvard Universitry
Press.
YOSHIMOTO, C.M. 1990. A review of the Genera of New World
Mymaridae (Hymenoptera: Chalcidoidea). Flora y Fauna Handbook
No. 7, 1-165 pp, Sandhill Crane Press.
YU, D.S. Y K. HORSTMANN. 1997a. A Catalogue of World Ichneumonidae
(Hymenoptera). Part I. Mem. Amer. Entomol. Soc., 58(1): 1-763.
YU, D.S. Y K. HORSTMANN. 1997b. A Catalogue of World Ichneumonidae
(Hymenoptera). Part II. Mem. Amer. Entomol. Soc., 58(2): 764-1558.
Anexo
I
Relación
de superfamilias, familias y subfamilias de
himenópteros de la región Neotropical
Al
frente de cada taxón se relaciona:
Estatus filogenético: Monofilético; Parafilético;
Sin información).
Gén/Sp: Números de géneros y especies
conocidos o estimados
Claves: disponibilidad de claves para género (G) o
especies (E)
Monografías y/o Catálogos para toda la región
o locales.
Subir referencia Anexo I
|
XYELOIDEA
Estatus filogenético: Parafilético - Gén/Sp:
1 - 1 - Claves: E - Monografías: Smith, 1988
C: Smith, 1978
Xyelidae
Estatus filogenético: Parafilético - Gén/Sp:
1 - 1 - Claves: G - Monografías: Smith, 1988
C: Smith, 1978
TENTHREDINOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
115 - 1.027 - Monografías: Smith, en prep.
Argidae
Estatus filogenético: Monofilético - Gén/Sp:
32 - 356 - Claves: E - Monografías: Smith, 1992
Arginae - Gén/Sp: 2 - 82 - Claves: E - Monografías:
Smith, 1992
Atomacerinae - Gén/Sp: 1 - 33 - Claves: E - Monografías:
Smith, 1992
Erigleninae - Gén/Sp: 4 - 41 - Claves: E - Monografías:
Smith, 1992
Dielocerinae - Gén/Sp: 6 - 31 - Claves: E - Monografías:
Smith, 1992
Sterictiphorinae - Gén/Sp: 19 - 169 - Claves:
E - Monografías: Smith, 1992
Cimbicidae
Estatus filogenético: Monofilético - Gén/Sp:
5 - 9 - Claves: E - Monografías: Smith, 1988
Pachylostictinae - Gén/Sp: 5 - 9 - Claves: E
- Monografías: Smith, 1988
Diprionidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 13 - Claves: E - Monografías: Smith, 1988
Monocteninae - Gén/Sp: 1 - 1 - Claves: E - Monografías:
Smith, 1988
Diprioninae - Gén/Sp: 2 - 12 - Claves: E - Monografías:
Smith, 1988
Pergidae
Estatus filogenético: Sin información.
- Gén/Sp: 32 - 256 - Claves: E - Monografías:
Smith, 1990
Acordulecerinae - Gén/Sp: 11 - 89 - Claves: E
- Monografías: Smith, 1990
Conocoxinae - Gén/Sp: 3 - 5 - Claves: E - Monografías:
Smith, 1990
Loboceratinae - Gén/Sp: 3 - 30 - Claves: E -
Monografías: Smith, 1990
Parasyzygoniinae - Gén/Sp: 1 - 2 - Claves: E
- Monografías: Smith, 1990
Pergulinae - Gén/Sp: 1 - 15 - Claves: E - Monografías:
Smith, 1990
Perreyinae - Gén/Sp: 6 - 81 - Claves: E - Monografías:
Smith, 1990
Philomastiginae - Gén/Sp: 2 - 2 - Claves: E -
Monografías: Smith,1990
Syzygoniinae - Gén/Sp: 5 - 32 - Claves: E - Monografías:
Smith, 1990
Tenthredinidae
Estatus filogenético: Sin información.-
43 - 393 - Claves: G - Monografías: Smith, en
prep.
Allantinae - Gén/Sp: 5 - 40 - Claves: E - Monografías:
Malaise, 1963 M: Smith, en prep.
Blennocampinae - Gén/Sp: 12 - 120 - Claves: E
- Monografías: Smith, en prep.
Heterarthrinae - Gén/Sp: 3 - 7 - Claves: E -
Monografías: Smith, 1973
Nematinae - Gén/Sp: 2 - 5 - Claves: E - Monografías:
Smith, en prep.
Selandriinae - Gén/Sp: 20 - 220 - Claves: E -
Monografías: Malaise, 1963 M: Smith, en prep.
Tenthredininae - Gén/Sp: 1 - 1 - Claves: E -
Monografías: Smith, en prep.
MEGALODONTOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
1 - 4 - Claves: E - Monografías: Smith, 1988
Pamphilidae
Estatus filogenético: Monofilético - 1
- 4 - Claves: E - Monografías: Smith, 1988
Cephalciinae - Gén/Sp: 1 - 4 - Claves: E - Monografías:
Smith, 1988
CEPHOIDEA
Gén/Sp: 1 - 1 - Claves: E - Monografías:
Smith, 1988
Cephidae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 1 - Claves: E - Monografías: Smith, 1988
Cephinae - Gén/Sp: 1 - 1 - Claves: E - Monografías:
Smith, 1988
SIRICOIDEA
Estatus filogenético: Parafilético - Gén/Sp:
10 - 27 - Claves: E - Monografías: Smith, 1988
Siricidae
Estatus filogenético: Monofilético - Gén/Sp:
6 - 10 - Claves: E - Monografías: Smith, 1988
Siricinae - Gén/Sp: 4 - 7 - Claves: E - Monografías:
Smith, 1988
Tremecinae - Gén/Sp: 2 - 3 - Claves: E - Monografías:
Smith, 1988
Xiphydriidae
Estatus filogenético: Monofilético - Gén/Sp:
4 - 17 - Claves: E - Monografías: Smith, 1988
Derecyrtinae - Gén/Sp: 3 - 16 - Claves: E - Monografías:
Smith, 1988
Xiphydriinae - Gén/Sp: 1 - 1 - Claves: E - Monografías:
Smith, 1988
ORUSSOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
5 - 13 - Claves: E - Monografías: Smith, 1988
Orussidae
Estatus filogenético: Monofilético - Gén/Sp:
5 - 13 - Claves: E - Monografías: Smith, 1988
STEPHANOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
2 - 27 - Claves: G - Monografías: Elliot, 1922
Stephanidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 27 - Claves: G - Monografías: Elliot, 1922;
Aguiar, 1998
TRIGONALOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
8 - 29 - Gén/Sp: Monografías y Claves:
Carmean y Kimsey, 1998
Trigonalidae
Estatus filogenético: Monofilético - Gén/Sp:
8 - 29 - Claves: G - Monografías y Claves: Carmean
y Kimsey,1998
Orthogonalinae - Estatus filogenético: Monofilético
- 1 - 1 - Claves: G - Monografías y Claves: Carmean
y Kimsey, 1998
Trigonalinae - Estatus filogenético: Monofilético
- Gén/Sp: 7 - 28 - Claves: G - Monografías
y Claves: Carmean y Kimsey, 1998
ICHNEUMONOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
694 - 4.637 - Gén/Sp: Goulet y Huber 1993
Braconidae
- Estatus filogenético: Monofilético -
Gén/Sp: 274 - 1.500 - Claves: G - Monografías:
Wharton et al., 1997; Campos (com. pers.)
Adeliinae - Gén/Sp: 1 - 2 - Claves: G - Monografías:
Wharton et al., 1998
Agathidinae - Gén/Sp: 18 - 800 - Claves: G -
Monografías: Wharton et al., 1998
Alysiinae - Gén/Sp: 19 - 250 - Claves: G - Monografías:
Wharton et al., 1998
Aphidiinae - Gén/Sp: 7 - 100 - Claves: G - Monografías:
Wharton et al., 1998
Betylobraconinae - Gén/Sp: 4 - 10 - Claves: G
- Monografías: Wharton et al., 1998
Braconinae - Gén/Sp: 24 - 800 - Claves: G - Monografías:
Wharton et al., 1998
Cardiochilinae - Gén/Sp: 7 - 30 - Claves: G -
Monografías: Wharton et al., 1998
Cenocoelinae - Gén/Sp: 4 - 10 - Claves: G - Monografías:
Wharton et al., 1998
Cheloninae - Gén/Sp: 8 - 120 - Claves: G - Monografías:
Wharton et al., 1998
Doryctinae - Gén/Sp: 65 - 700 - Claves: G - Monografías:
Wharton et al., 1998
Euphorinae - Gén/Sp: 25 - 250 - Claves: G - Monografías:
Wharton et al., 1998; Shaw, 1988
Gnamptodontinae - Gén/Sp: 2 - 15 - Claves: G
- Monografías: Wharton et al., 1998
Helconinae - Gén/Sp: 7 - 500 - Claves: G - Monografías:
Wharton et al., 1998
Homolobinae - Gén/Sp: 2 - 15 - Claves: G - Monografías:
Wharton et al., 1998
Hormiinae - Gén/Sp: 8 - 150 - Claves: G - Monografías:
Wharton et al., 1998
Ichneutinae - Gén/Sp: 8 - 30 - Claves: G - Monografías:
Wharton et al., 1998
Macrocentrinae - Gén/Sp: 4 - 50 - Claves: G -
Monografías: Wharton et al., 1998
Mendesellinae - Gén/Sp: 2 - 10 - Claves: G -
Monografías: Wharton et al., 1998
Meteorideinae - Gén/Sp: 1 - 3 - Claves: G - Monografías:
Wharton et al., 1998
Meteorinae - Gén/Sp: 2 - 80 - Claves: G - Monografías:
Wharton et al., 1998
Microgastrinae - Gén/Sp: 36 - 500 - Claves: G
- Monografías: Wharton et al., 1998
Miracinae - Gén/Sp: 1 - 10 - Claves: G - Monografías:
Wharton et al., 1998
Opiinae - Gén/Sp: 2 - 70 - Claves: G - Monografías:
Wharton et al., 1998
Orgilinae - Gén/Sp: 2 - 65 - Claves: G - Monografías:
Wharton et al., 1998
Rogadinae - Gén/Sp: 8 - 100 - Claves: G - Monografías:
Wharton et al., 1998
Sigalphinae - Gén/Sp: 2 - 5 - Claves: G - Monografías:
Wharton et al., 1998
Ypsistocerinae - Gén/Sp: 3 - 5 - Claves: G -
Monografías: Wharton et al., 1998
Ichneumonidae
Estatus filogenético: Monofilético - Gén/Sp:
420 - 3.137 - Claves: G - Monografías: Townes,
1969 - 1971 Claves: Townes y Townes, 1966; Yu y
Horsmann, 1997a, b; Palacio (com. pers.)
Acaenitinae - Gén/Sp: 1 - 2 - Wahl y Palacio,
en prep.
Anomaloninae - Gén/Sp: 19 - 1 - Wahl y Palacio,
en prep.
Banchinae - Gén/Sp: 24 - 82/1501 - Wahl y Palacio,
en prep.
Camplopeginae - Gén/Sp: 20 - 170 - Wahl y Palacio,
en prep.
Cremastinae - Gén/Sp: 12 - 67 - Wahl y Palacio,
en prep.
Cryptinae - Gén/Sp: 130 - 819 - Wahl y Palacio,
en prep.
Ctenopelmatelinae - Gén/Sp: 17 - 65 - Wahl y
Palacio, en prep.
Cyllocerinae - Gén/Sp: 1 - 6 - Wahl y Palacio,
en prep.
Diplazontinae - Gén/Sp: 8 - 76 - Wahl y Palacio,
en prep.
Eucerotinae - Gén/Sp: 1 - 1 - Wahl y Palacio,
en prep.
Ichneumoninae - Gén/Sp: 72 - 423 - Wahl y Palacio,
en prep.
Labeninae - Gén/Sp: 5 - 39 - Wahl y Palacio,
en prep.
Lycorininae - Gén/Sp: 1 - 9 - Wahl y Palacio,
en prep.
Mesochorinae - Gén/Sp: 5 - 277 - Wahl y Palacio,
en prep.
Metopiinae - Gén/Sp: 10 - 24/701 - Wahl y Palacio,
en prep.
Neorhacodinae - Gén/Sp: 1 - 17 - Wahl y Palacio,
en prep.
Ophioninae - Gén/Sp: 13 - 212 - Wahl y Palacio,
en prep.
Orthocentrinae - Gén/Sp: 16 - 12/751 - Wahl y
Palacio, en prep.
Oxytorinae - Gén/Sp: 1 - 1 - Wahl y Palacio,
en prep.
Phrudinae - Gén/Sp: 1 - 2 - Wahl y Palacio, en
prep.
Pimplinae - Gén/Sp: 29 - 297/3363 - Wahl y Palacio,
en prep.
Poemeniinae - Gén/Sp: 2 - 3 - Wahl y Palacio,
en prep.
Rhyssinae - Gén/Sp: 2 - 45 - Wahl y Palacio,
en prep.
Stilbopinae - Gén/Sp: 2 - 2 - Wahl y Palacio,
en prep.
Tatogastrinae - Gén/Sp: 1 - 1 - Wahl y Palacio,
en prep.
Tersilochinae - Gén/Sp: 5 - 10/351 - Wahl y Palacio,
en prep.
Tryphoninae - Gén/Sp: 14 - 100 - Wahl y Palacio,
en prep.
Xoridinae - Gén/Sp: 2 - 16 - Wahl y Palacio,
en prep.
CHRYSIDOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
78 - 1.177
Plumariidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 18 - Claves: E - Roig - Alsina, 1994
Scolebythidae
Gén/Sp: 2 - 3 - Claves: E - Evans, 1963; Azevedo,
1999
Bethylidae
Estatus filogenético: Monofilético - Gén/Sp:
26 - 437 - Claves: E - Evans, 1964; Hawkins y Gordh,
1986; C: Gordh y Moczar, 1990; Vargas y Terayama, en
prep.
Bethylinae - Gén/Sp: 3 - 46
Epyrinae - Gén/Sp: 18 - 195
Pristocerinae - Gén/Sp: 5 - 196
Chrysididae
Estatus filogenético: Monofilético - Gén/Sp:
24 - 278 - Monografías y Claves: Kimsey y Bohart,
1990
Amiseginae - Gén/Sp: 5 - 38
Cleptinae - Gén/Sp: 2 - 16
Chrysidinae - Gén/Sp: 17 - 224
Sclerogibbidae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 2 - Claves: E - Richards, 1939; Argaman, 1988
Dryinidae
Estatus filogenético: Monofilético - Gén/Sp:
21 - 430 - Claves: E - Monografías: Olmi, 1984,
1989, 1995a
Anteoninae - Gén/Sp: 4 - 117
Aphelopinae - Gén/Sp: 2 - 18
Apodryininae - Gén/Sp: 1 - 1
Bocchinae - Gén/Sp: 1 - 4
Dryininae - Gén/Sp: 2 - 64
Gonatopodinae - Gén/Sp: 11 - 80
Embolemidae
Estatus filogenético: Monofilético - Gén/Sp:
2 - 10 - Claves: E - Monografías: Olmi, 1995b;
Amarante et al., 1999
APOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
352 - 5.000 - Michener 1997, 2000
"spheciformes" - Estatus filogenético:
Parafilético - Gén/Sp: 141 - 1.630 - Monografías
y Catálogos: Bohart y Menke, 1976 M: Menke y
Fernández, 1996
Ampulicidae
Gén/Sp: 3 - 20 - Monografías y Claves:
Bohart y Menke, 1976
Sphecidae
Gén/Sp: 14 - 159 - Monografías y Claves:
Bohart y Menke, 1976
Crabronidae
Gén/Sp: 124 - 1.451 - Monografías y Claves:
Bohart y Menke, 1976
Pemphredoninae - Gén/Sp: 17 - 158 - Monografías
y Catálogos Bohart y Menke, 1976
Astatinae - Gén/Sp: 4 - 22 - Monografías
y Catálogos: Bohart y Menke, 1976
Crabroninae - Gén/Sp: 45 - 751 - Monografías
y Catálogos: Bohart y Menke, 1976
Bembecinae - Gén/Sp: 52 - 303 - Monografías
y Catálogos: Bohart y Menke, 1976
Philanthinae - Gén/Sp: 6 - 215 - Monografías
y Catálogos: Bohart y Menke, 1976
‘apiformes’ - Estatus filogenético:
Monofilético - Gén/Sp: 211 - 3.370 - Michener,
2000
Colletidae
Estatus filogenético: Monofilético - Gén/Sp:
22 - 442 - Catálogos: Michener, 1997; M: Michener,
2000
Colletinae - Gén/Sp: 8 - 238 - Monografías:
Michener, 2000
Diphaglossinae - Gén/Sp: 9 - 80 - Monografías:
Michener, 2000
Hylaeinae - Gén/Sp: 1 - 50 - Monografías:
Michener, 2000
Xeromelissinae - Gén/Sp: 4 - 74 - Monografías:
Michener, 2000
Andrenidae
Estatus filogenético: Monofilético - Gén/Sp:
23 - 238 - Monografías: Michener, 2000
Alocandreninae - Gén/Sp: 1 - 1 - Monografías:
Michener, 2000
Andreninae - Gén/Sp: 3 - 5 - Monografías:
Michener, 2000
Oxaeinae - Gén/Sp: 2 - 12 - Monografías:
Michener, 2000
Panurginae - Gén/Sp: 17 - 220 - Monografías:
Michener, 2000
Halictidae
Estatus filogenético: Monofilético - Gén/Sp:
43 - +800 - Monografías: Moure y Hurd, 1987;
Michener, 2000
Halictinae - Gén/Sp: 40 - +788 - Monografías:
Michener, 2000
Nomiinae - Gén/Sp: 1 - 1 - Monografías:
Michener, 2000
Rophitinae - Gén/Sp: 2 - 2 - Monografías:
Michener, 2000
Megachilidae
Gén/Sp: 19 - +350 - Monografías: Michener,
2000
Fideliinae - Gén/Sp: 1 - 1 - Monografías:
Michener, 2000
Megachilinae - Gén/Sp: 18 - +349 - Monografías:
Michener, 2000
Apidae - Gén/Sp: 104 - 1.500 - Monografías:
Michener, 2000
Apinae - Gén/Sp: 90 - 1.176 - Monografías:
Michener, 2000
Nomadinae - Gén/Sp: 13 - 191 - Monografías:
Michener, 2000
Xylocopinae - Gén/Sp: 3 - 140 - Monografías:
Michener, 2000
VESPOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
373 - 6.309
Sierolomorphidae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 5 - Claves: E - Monografías: Evans, 1961
Rhopalosomatidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 20 - Claves: E - Monografías: Townes, 1977a
Bradynobaenidae
Estatus filogenético: Monofilético - Gén/Sp:
4 - 14 - Claves: G - Monografías: Genise, 1986
Bradynobaeninae - Gén/Sp: 1 - 6
Typhoctinae - Gén/Sp: 3 - 8 - Quintero y Rodríguez,
1990
Formicidae
Estatus filogenético: Monofilético - Gén/Sp:
120 - 2.365 - Claves: G - Monografías: Bolton,
1994; Catálogos: Kempf, 1972; Bolton, 1995
Ponerinae - Estatus filogenético: Parafilético
- 24 - 348 - Claves: G -
Cerapachyinae - Estatus filogenético: Monofilético
- 4 - 25 - Claves: E -
Ecitoninae - Estatus filogenético: Monofilético
- 5 - 129 - Claves: E - Monografías: Watkins,
1976
Leptanilloidinae - Estatus filogenético: Monofilético
- 2 - 3 - Claves: E - Monografías: Brandao et
al., 1999
Pseudomyrmecinae - Estatus filogenético: Monofilético
- 2 - 115 - Claves: E - Monografías: Ward, 1990
Myrmicinae - Estatus filogenético: Monofilético
- 68 - 1.113 - Claves: G -
Dolichoderinae - Estatus filogenético: Monofilético
- 8 - 209 - Claves: G - Monografías: Shattuck,
1992 C: Shattuck, 1994
Formicinae - Estatus filogenético: Monofilético
- 9 - 422 - Claves: G -
Vespidae
Estatus filogenético: Monofilético - 79
- 1.330 - Claves: G -
Masarinae - Estatus filogenético: Monofilético
- 4 - 20 - Claves: E - Monografías: Richards,
1962; Sarmiento, 1999
Eumeninae - Estatus filogenético: Monofilético
- 50 - +800 - Claves: G - Carpenter, 1986b, en prep.;
Sarmiento, 1999
Polistinae - Estatus filogenético: Monofilético
- 24 - +500 - Claves: E - Monografías: Richards,
1978
Vespinae - Estatus filogenético: Monofilético
- 1 - 4 - Claves: E -
Scoliidae
Estatus filogenético: Monofilético - 2
- 40 - Claves: E - Monografías: Bradley, 1945
Scoliinae - Estatus filogenético: Monofilético
- 2 - 40 - Claves: E -
Tiphiidae
Estatus filogenético: Monofilético - Gén/Sp:
35 - 228 - Monografías: Kimsey, 1991
Anthoboscinae - Estatus filogenético: Monofilético
- 4 - 16 - Claves: E - Monografías: Genise, 1984
Brachystidinae - Estatus filogenético: Monofilético
- 1 - 2 - Claves: E - Monografías: Wasbauer,
1968
Methocinae - Estatus filogenético: Monofilético
- 1 - 8? - Claves: G - Monografías: Krombein,
1979
Myzininae - Estatus filogenético: Monofilético
- 2 - 10? - Claves: G - Monografías: Krombein,
1937
Tiphiinae - Estatus filogenético: Monofilético
- 4 - 92 - Claves: E -Monografías: Allen, 1972;
Kimsey, 1993
Thynninae - Estatus filogenético: Monofilético
- 23 - 100? - Claves: G - Monografías: Kimsey,
1992
Pompilidae
Estatus filogenético: Monofilético - Gén/Sp:
57 - 800 - Claves: G - Monografías: Banks, 1946,
1947; Evans, 1966
Ceropalinae - Estatus filogenético: Monofilético
- 2 - 18 -
Notocyphinae - Estatus filogenético: Monofilético
- 1 - 34 -
Pepsinae - Estatus filogenético: Monofilético
- 24 - 460 -
Pompilinae - Estatus filogenético: Monofilético
- 30 - 275 -
Mutillidae
Estatus filogenético: Monofilético - Gén/Sp:
69 - 1.500 - Catálogos: Nonvellier, 1990
Mutillinae - Estatus filogenético: Monofilético
- 6 - 400 -
Sphaerophtalminae - Estatus filogenético: Monofilético
- 63 - 1.100 -
Sapygidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 7 - Claves: G - Monografías: Pate, 1947
Sapyginae - Gén/Sp: 3 - 7 - Claves: G - Monografías:
Pate, 1947
CYNIPOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
+60 - +800 - Gén/Sp: Monografías: Weld,
1952
Liopteridae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 57 - Gén/Sp: Monografías: Ronquist,
1995b
Figitidae
Estatus filogenético: Monofilético - Gén/Sp:
44 - 225 - Gén/Sp: Monografías: Weld,
1952
Anacharitinae - Gén/Sp: 5 - 10 -
Aspicerinae - Gén/Sp: 3? - 15 -
Figitinae - Gén/Sp: 36? - +200 -
Cynipidae
Estatus filogenético: Monofilético - Gén/Sp:
10 - 15 - Gén/Sp: Monografías: Weld, 1952
Cynipinae - Gén/Sp: 10 - 15 -
PROCTOTRUPOIDEA
Estatus filogenético: Parafilético - Gén/Sp:
94 - 334 - Catálogos: Johnson, 1992
Monomachidae
Estatus filogenético: Monofilético - Gén/Sp:
2 - 10 - Claves: G -
Heloridae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 2 - Claves: E - Monografías: Townes, 1977b
Pelecinidae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 1 - Claves: E - Monografías: Goulet y Huber,
1993
Proctotrupidae
Estatus filogenético: Monofilético - Gén/Sp:
8 - 93 - Claves: E - Monografías: Townes y Townes,
1981
Austroserphinae - Gén/Sp: 1 - 1 -
Proctotrupinae - Gén/Sp: 7 - 92 -
Diapriidae
Estatus filogenético: Monofilético - Gén/Sp:
84 - 229 -
Ambositrinae - Gén/Sp: 4 - 8 -
Belytinae - Gén/Sp: 21 - 26 -
Diapriinae - Gén/Sp: 58 - 180 - Claves: G - Masner
y García, en prep.
Ismarinae - Gén/Sp: 1 - 15 - Claves: E - Monografías:
Masner, 1976b
PLATYGASTROIDEA
Estatus filogenético: Monofilético - Gén/Sp:
82 - 434 - C: Johnson, 1992; Vlug, 1995
Scelionidae
Estatus filogenético: Monofilético - Gén/Sp:
56 - 334 - Monografías: Masner, 1976a, 1980
Scelioninae - Gén/Sp: 44 - 227 -
Teleasinae - Gén/Sp: 6 - 16 -
Telenominae - Gén/Sp: 6 - 91 -
Platygastridae
Estatus filogenético: Monofilético - Gén/Sp:
26 - +100 - Monografías: Masner y Huggert, 1989
Platygastrinae - Gén/Sp: +20 -
Sceliotrachelinae - Gén/Sp: 8 -
MYMAROMMATOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
1 - 1
Mymarommatidae
Estatus filogenético: Monofilético - Gén/Sp:
1 - 1 - Claves: G -
CHALCIDOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
625 - 3.300 - C: De Santis, 1979, 1983, 1989
Chalcididae
Estatus filogenético: Parafilético - Gén/Sp:
21 - 217 - Monografías: Delvare y Boucek, 1992;
Boucek, 1992
Chalcidinae - Gén/Sp: 13 - 199 -
Dirhininae - Gén/Sp: 1 - 8 -
Epitraninae - Gén/Sp: 1 - 3 -
Haltichellinae - Gén/Sp: 11 - 7 -
Leucospidae
Estatus filogenético: Monofilético - Gén/Sp:
2 - 34 - Monografías: Boucek, 1974
Eurytomidae
Estatus filogenético: Monofilético - Gén/Sp:
21 - 125 -
Eurytominae - Gén/Sp: 16 - 74 -
Heimbrinae - Gén/Sp: 2 - 5 -
Rileyinae - Gén/Sp: 3 - 31 -
Pteromalidae
Estatus filogenético: Parafilético - Gén/Sp:
84 - 242 -
Asaphinae - Gén/Sp: 1 - 2 -
Ceinae - Gén/Sp: 1 - 2 -
Cerocephalinae - Gén/Sp: 4 - 4 -
Cleonyminae - Gén/Sp: 3 - 26 -
Colotrechninae - Gén/Sp: 4 - 8 -
Diparinae - Gén/Sp: 1 - 27 -
Dvaliniinae - Gén/Sp: 4 - 5 - Gén/Sp:
Erotolepsiinae - Gén/Sp: 1 - 1 -
Eunotinae - Gén/Sp: 4 - 4 -
Eutrichosomatinae - Gén/Sp: 2 - 3 -
Herbertiinae - Gén/Sp: 1 - 1 -
Leptofoeninae - Gén/Sp: 1 - 4 -
Miscogasterinae - Gén/Sp: 19 - 56 -
Ormocerinae - Gén/Sp: 3 - 3 -
Pireninae - Gén/Sp: 4 - 4 -
Pteromalinae - Estatus filogenético: Parafilético
- 33 - 81 -
Spalangiinae - Gén/Sp: 1 - 11 -
Agaonidae
Estatus filogenético: Monofilético - Gén/Sp:
11 - 133 - Gén/Sp: Monografías: Ramirez,
1970
Agaoninae - Gén/Sp: 2 - 40 -
Blastophaginae - Gén/Sp: 1 - 27 -
Epichrysomalinae - Gén/Sp: 4 - 9 -
Otitesellinae - Gén/Sp: 2 - 40 -
Sycophaginae - Gén/Sp: 1 - 12 -
Sycoryctinae - Gén/Sp: 1 - 5 -
Torymidae
Gén/Sp: 11 - 60 -
Megastigminae - Gén/Sp: 1 - 2 -
Monodontomerinae - Gén/Sp: 5 - 15 -
Podagrioninae - Gén/Sp: 1 - 12 -
Toryminae - Gén/Sp: 4 - 29 -
Ormyridae
Gén/Sp: 1 - 2 - Monografías: Hanson, 1992
Perilampidae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 22 - Monografías: Heraty y Darling, 1984
Eucharitidae
Estatus filogenético: Parafilético - Gén/Sp:
11 - 40 - Monografías: Heraty y Darling, 1984
Eucharitinae - Gén/Sp: 10 - 28 -
Oraseminae - Gén/Sp: 1 - 12 -
Eupelmidae
- Gén/Sp: Gén/Sp: 15 - 85 - Monografías:
Gibson, 1989
Calosotinae - Gén/Sp: 2 - 2 -
Eupelminae - Gén/Sp: 11 - 72 -
Neanastatinae - Gén/Sp: 2 - 11 -
Tanaostigmatidae
Estatus filogenético: Monofilético - Gén/Sp:
6 - 60 - Monografías: La Salle, 1987
Encyrtidae
Gén/Sp: 147 - +500 - Monografías: Noyes,
1980
Aphelinidae
Estatus filogenético: Parafilético - Gén/Sp:
22 - 119 -
Aphelininae - Gén/Sp: 6 - 37 -
Azotinae - Gén/Sp: 1 - 1 -
Calesinae - Gén/Sp: 1 - 1 -
Pterotricinae - Gén/Sp: 4 - 12 -
Signiphoridae
Gén/Sp: 2 - 28 -
Signiphorinae - Gén/Sp: 2 - 28 -
Thysaninae - Gén/Sp: 3 - 4 -
Tetracampidae
Gén/Sp: 1 - 1 -
Rotoitidae
Gén/Sp: 1 - 1 - Monografías: Boucek y
Noyes, 1987
Eulophidae
Gén/Sp: 55 - 227 -
Elachertinae - Gén/Sp: 12 - 37 -
Entedoninae - Gén/Sp: 23 - 85 -
Eudorinae - Gén/Sp: 6 - 11 -
Eulophinae - Gén/Sp: 6 - 8 -
Tetrastichinae - Gén/Sp: 8 - 86 -
Elasmidae
Gén/Sp: 1 - 13 -
Trichogrammatidae
Estatus filogenético: Monofilético - Gén/Sp:
20 - 55 -
Mymaridae
Estatus filogenético: Monofilético - Gén/Sp:
40 - 177 - Monografías: Yoshimoto, 1990; Huber,
1986
MEGALYROIDEA
Estatus filogenético: Monofilético - Gén/Sp:
3 - 3 - Monografías: Shaw, 1990
Megalyridae
Estatus filogenético: Monofilético - Gén/Sp:
3 - 3 - Shaw, 1990
CERAPHRONOIDEA
Estatus filogenético: Monofilético - Gén/Sp:
12 - +90 - Monografías: Dessart y Cancemi, 1986
Megaspilidae
Estatus filogenético: Monofilético - Gén/Sp:
6 - +40 - Monografías: Dessart y Cancemi, 1986
Lagynodinae - Gén/Sp: 1 - +10 -
Megaspilinae - Gén/Sp: 5 - +30 -
Ceraphronidae
Estatus filogenético: Monofilético - Gén/Sp:
6 - +50 - Monografías: Dessart y Cancemi, 1986
EVANIOIDEA
Estatus filogenético: Parafilético - Gén/Sp:
10 - +180 -
Aulacidae
Estatus filogenético: Monofilético - Gén/Sp:
2 - 50 - Smith, en prep.
Gasteruptiidae
Gén/Sp: 2 - +30 -
Evaniidae
Gén/Sp: 6 - +100
|
Subir referencia Anexo
I
|
Tabla
I
Caracteres
sinapomórficos o potencialmente sinapomórficos
propuestos para Hymenoptera (recopilados por Vilhelmsen,
1997b)
Parte
flexionada del clípeo esclerotizada.
Presencia de un complejo labiomaxilar.
Fusión entre laterocervical y pre - episterno.
Músculos posteriores notocoxales ausentes y meron
no demarcado sobre la coxa.
Trocantelo (prefémur) presente.
Espolón tibial anterior apical modificado (p.e.
en limpiador de antena).
Venas anales del ala anterior no alcanzan el margen
del ala.
Presencia de hamuli sobre el margen anterior del ala
posterior.
Presencia de brazos anteriores mesofurcales.
Músculo mesoescutelar metanotal unido al metanoto.
Presencia de dos músculos mesofurcal laterofragmales.
Primer tergo abdominal fuertemente asociado con el metapostnoto.
Tergos abdominales sobrelapan los esternos y rodean
los espiráculos abdominales.
Anillo basal y volsela presentes en genitalia del macho
Presencia de una articulación basal en el ovipositor.
Ojo en larvas (cuando están presentes) con una
cornea única sobrepuestas a varias omatidias.
Determinación de sexo haplodiploide (generalmente
hembras diploides y machos haploides).
|
Subir referencia Tabla
I
Tabla
II
Comparación
de superfamilias: número de géneros y
número
de especies descritos por superfamilia.
Los datos de Cynipoidea y Ceraphronoidea son tentativos.
|
Superfamilias
|
Número
géneros
|
Número
especies
|
| Xyeloidea |
1
|
1
|
| Tenthredinoidea |
115
|
1027
|
| Megalodontoidea |
1
|
4
|
| Cephoidea |
1
|
1
|
| Siricoidea |
10
|
27
|
| Orussoidea |
5
|
13
|
| Stephanoidea |
2
|
27
|
| Trigonaloidea |
8
|
29
|
| Ichneumonoidea |
694
|
4637
|
| Chrysidoidea |
78
|
1177
|
| Apoidea |
352
|
5000
|
| Vespoidea |
373
|
6309
|
| Cynipoidea |
60
|
800
|
| Proctotrupoidea |
94
|
334
|
| Platygastroidea |
82
|
434
|
| Mymarommatoidea |
1
|
1
|
| Chalcidoidea |
625
|
3300
|
| Megalyroidea |
3
|
3
|
| Ceraphronoidea |
12
|
90
|
| Evanioidea |
10
|
180
|
|
Totales
.
|
2527
|
23394
|
|
Subir referencia Tabla II
|
.... |
