Arácnidos Fósiles (con exclusión de
Arañas y Escorpiones)
Jason A. Dunlop
Introducción
Los arácnidos son uno de los grupos de
artrópodos actuales más importantes. Los
fósiles de arácnidos son escasos en
comparación con los de otros grupos
animales, pero los conocidos pueden ser muy
importantes en la determinación de las
relaciones evolutivas entre los grupos de
arácnidos. El registro fósil incluye órdenes
ahora extinguidos, es decir, "dinosaurios"
del mundo de las arañas. Entre los resúmenes
recientes de arácnidos fósiles y su
importancia están Shear & Kukalova-Peck
(1990) y Selden (1993a, b) presenta una
panorámica del material fósil. El material
fósil de arañas y escorpiones se revisa en
otro lugar de este mismo volumen. En el
presente trabajo se hace una revisión
cronológica del material fósil de los otros
órdenes de arácnidos, y se comentan el
momento de aparación y la presencia en
diversos períodos geológicos de estos
órdenes (resumidos en la Fig. 1), así como
la aplicación del registro fósil a la
filogenia de los arácnidos.

Figura 1:
Amplitud estratigráfica de los
órdenes de arácnidos. Adaptado de Selden
(1993a, b) y actualizado para incluir
información inédita. Las barras gruesas
indican que se conoce la existencia de
fósiles, las líneas finas son lagunas en
el registro fósil.
Períodos geológicos
El tiempo geológico
se clasifica en una serie de divisiones
reconocidas a nivel global. El Fanerozoico -
que comprende aproximadamente los
seiscientos millones de años en los que los
animales multicelulares son el grupo
dominante-, se divide en tres eras, el
Paleozoico (vida antigua), Mesozoico (vida
media) y Cenozoico (vida reciente). Cada una
de estas eras se subdivide además en
sistemas. En este capítulo vamos a seguir la
pista del registro fósil y evolución de los
arácnidos y grupos relacionados a través de
estos sistemas geológicos. Las edades
geológicas se dan en millones de años antes
de la actualidad (Ma), y siguen la división
estratigráfica global de Cowie & Bassett
(1989).
EL PALEOZOICO
El Paleozoico es
quizás la época más interesante para la
evolución de los arácnidos. Durante el
Paleozoico evolucionaron los quelicerados, a
partir de un antepasado aún desconocido, y
además evolucionaron los Arachnida a partir
de los Chelicerata. Ahora parece probable
que al menos dos grupos de arácnidos, por un
lado los escorpiones y por otro lado el
resto de los arácnidos, llegasen a tierra
firme de forma independiente. Durante el
Paleozoico parecen haber quedado definidos
todos los órdenes de arácnidos actuales (no
hay fósiles paleozoicos de Palpigradi ni
Schizomida, pero puede que hayan estado
presentes, aunque no tienen facilidad para
conservarse en estado fósil). En el
Paleozoico estaban presentes también tres
órdenes ahora desaparecidos, Trigonotarbida,
Phalangiotarbida y Haptopoda. Hay un cambio
claro en la fauna de arácnidos al final del
Paleozoico, con la extinción de estos tres
órdenes y a continuación una posible
radiación de las arañas.
Cámbrico (570-510 Ma)
No se conocen arácnidos del Cámbrico ni del
Ordovícico, pero el origen de los
Chelicerata, el grupo al que pertenecen los
arácnidos, puede seguirse hasta el Cámbrico.
Los quelicerados evolucionaron probablemente
en el Cámbrico como parte de la llamada
"explosión cámbrica" (Gould, 1989), en la
que aparecieron muchos artrópodos nuevos. Se
suele aceptar que los Chelicerata pertenecen
a un amplio grupo de artrópodos llamados los
Arachnomorpha (o Arachnata), grupo que
incluye a quelicerados, trilobites y una
serie de fósiles "problemáticos". Algunos de
estos fósiles problemáticos, sobre todo del
famoso yacimiento de Burgess Shale, en
Canadá (≈530 Ma), se
parecen a los xifosuros (cacerolas de las
Molucas) y en principio se pensó que eran
quelicerados (Walcott, 1912; Størmer, 1944).
Estos fósiles, denominados "merostomoides",
fueron revisados por Harry Whittington y sus
colegas (e.g. Bruton & Whittington, 1983),
quienes sugirieron que los merostomoides no
eran quelicerados, ni siquiera antepasados
de los quelicerados, sino parte de un amplio
grupo de artrópodos no emparentados (ideas
resumidas por Gould (1989)). Recientes
análisis cladísticos de estos artrópodos
problemáticos (e.g. Wills et al., 1995)
sugieren que estos grupos difíciles tienen
algún parentesco. Estos autores proponen que
los Arachnomorpha forman un clado de
artrópodos que contarían con los Chelicerata
como representantes vivos. De todas formas,
las relaciones de los quelicerados con otros
fósiles aracnomorfos siguen estando claras.
Hay una serie de
fósiles cámbricos a los que se ha
clasificado como quelicerados. Un grupo de
fósiles parecidos a los xifosuros, los
aglaspídidos, ha estado estado incluido en
los Chelicerata durante muchos años (e.g.
Raasch, 1939). Los aglaspídidos fueron
excluidos de los Chelicerata por Briggs et
al. (1979), pero algunos autores (e.g.
Weygold & Paulus, 1979) los consideran grupo
hermano de los quelicerados. Sanctacaris,
descrito de Burgess Shale por Briggs &
Collins (1988) se ha considerado en general
como quelicerado, pero en estudios recientes
no aparece como grupo hermano de otros
quelicerados y parece ser otro fósil
problemático en lugar de un antepasado de
las arañas, como a veces se le ha
considerado. Como respuesta al problema de
la identificación de los quelicerados
primitivos, Dunlop & Selden (en prensa)
redefinen los Chelicerata como artrópodos
con un tubérculo ocular medio bien definido
y un opistosoma más o menos difeferenciado.
Estos autores, por consiguiente, limitan los
Chelicerata a los Xiphosura, Chasmataspida,
Eurypterida y Arachnida. Ninguno de estos
grupos se conoce todavía del Cambrico, y por
tanto los quelicerados claros más antiguos
son del siguiente sistema geológico, el
Ordovícico.
Ordovícico (510-438 Ma)
Los quelicerados más
antiguos son del Ordovícico. Un fósil
diminuto y mal conservado, Lemoneites,
descrito por Flower (1968) del Ordovícico de
Nuevo Méjico, parece ser nuestra mejor
imagen del aspecto que tenían los primeros
quelicerados (Dunlop & Selden, en prensa).
Un grupo extinguido, los casmatáspidos
fueron originalmente considerados como
sifosuro atípicos, y el fósil devónico
Diploaspis fue, incluso, propuesto como
pariente próximo del antepasado de los
arácnidos (Bergström, 1979). Sin embargo,
Dunlop & Selden (en prensa) consideran a los
casmatáspidos como simple grupo, aunque bien
diferenciado, dentro de los Chelicerata.
Un grupo mejor conocido
de quelicerados extinguidos son los
euriptéridos, o "escorpiones de mar". Este
diversificado grupo de artrópodos predadores
aparece en el Ordovícico inferior (≈470 Ma),
y se encuentra en todo el Paleozoico
(Selden, 1984). Algunos, como
Baltoeurypterus, se han conservado con gran
detalle (Holm, 1898). Los euriptéridos
incluyen algunos de los artrópodos más
grandes que se conocen, como el Pterygotus,
de dos metros de largo. La mayoría de los
autores ven en los euriptéridos el grupo
hermano de los arácnidos (Weygoldt & Paulus,
1979; Shultz, 1990). En libros de cierta
antigüedad los quelicerados se dividían en
Merostomata, acuáticos (xifosuros y
euriptéridos), y Arachnida, terrestres. Sin
embargo, si los euriptéridos están más
emparentados con los arácnidos que los
xifosuros, "Merostomata" deja de ser un
término válido para estos dos grupos (Kraus,
1976), y el término merostoma debería dejar
de usarse. No hay fósiles de arácnidos del
Ordovícico, aunque, si los arácnidos y los
euriptéridos son grupos hermanos cabe
esperar que los arácnidos evolucionasen en
el Ordovícico.
Silúrico (438-410 Ma)
Entre los
quelicerados en general, los euriptéridos
(escorpiones de mar) alcanzaron su máxima
diversidad y abundancia en el Silúrico
(Selden, 1984). En el Silúrico también
encontramos los primeros xifosuros claros
(ver Anderson & Selden, (en prensa) para una
revisión actualizada de los Xiphosura
paleozoicos y de su filogenia). Tanto
sifosuros como euriptéridos se interpretan
como acuáticos, y presumiblemente los
primeros arácnidos eran también acuáticos.
Los arácnidos más antiguos conocidos son
escorpiones acuáticos del Silúrico medio (≈425
Ma). El arácnido terrestre más antiguo que
se conoce pertenece al desaparecido orden de
los Trigonotarbida, de aspecto araña. Este
fósil (Fig. 2b), notable pese a su tamaño
diminuto (1'5 mm) y su mal estado de
conservación, es del Silúrico superior (≈414
Ma), y proviene de Shropshire, en
Inglaterra. El espécimen fue citado por
Jeram et al. (1990), y ha sido descrito
recientemente como Eotarbus por Dunlop
(1996b). Este fósil se ha conservado como un
trozo oscuro, carbonizado, de la cutícula
originaria. Fue estraído de la roca por
disolución de roca, rica en cutícula, en
ácido hidrofluorídrico, que destruye la
matriz rocosa y deja sólo los fragmentos de
artrópodos y plantas. Los mismos métodos se
usaron para preparar los fósiles de la
localidad devónica de Gilboa (ver más
adelante).
Los trigonotárbidos son
un grupo significativo en el Paleozoico.
Tienen el aspecto de arañas, pero carecen de
hileras productoras de seda, y se
caracterizan por tener en el opistosoma
terguitos divididos en placas medianas y
laterales. Los trigonotárbidos llegaban a 5
cm de longitud, y muchas formas tardías
tenían cuerpos muy tuberculados, que han
llevado a algunos autores a llamarlos
"arañas acorazadas". Durante el Paleozoico
los fósiles de trigonotárbidos son más
abundantes que los de arañas y que la
mayoría de los otros arácnidos (quizás con
excepción de los escorpiones). Aunque
sabemos muy poco de los primeros ácaros y
pseudoescorpiones, sí podemos decir que
entre las grandes arácnidos los
trigonotárbidos eran evidentemente un grupo
dominante. El trigonotárbido Eotarbus es uno
de los primeros animales terrestres que se
conocen. Pertenece a la fauna más antigua
conocida de artrópodos terrestres (Jeram et
al., 1990). El registro fósil de los
primeros artrópodos terrestres, y los
ecosistemas en que vivían, se revisan en
Shear & Kukalová-Peck (1990) y Shear (1991).
Devónico (410-355 Ma)
Los fósiles de
arácnidos del Devónico incluyen nuevos
ejemplos de los primeros ecosistemas
terrestres. Los mejor conservados son del
Devónico inferior (≈398
Ma) de Rhynie Chert, en Escocia. Estos
fósiles fueron descritos originalmente por
Hirst (1923), y en su mayor parte contienen
el trigonotárbido Palaeocharinus (Fig. 2a).
También se describió un ácaro, Protacarus,
aunque Dubinin (1962) reconoce una mayor
diversidad de ácaros en Rhynie Chert. Los
fósiles de Rhynie son tridimensionales, y
están tan bien conservados porque quedaron
atrapados en agua rica en silicio, que se
endureció formando un chert. Se ha
conservado bien cualquier parte del cuerpo
que estuviera cubierta por cutícula. En los
trigonotárbidos de Rhynie se encuentran los
ejemplos más antiguos conocidos de
filotráqueas (Claridge & Lyon, 1961).
También presentan un sistema de filtración
preoral (Dunlop, 1994a), lo que sugiere que
ingerían comida líquida como las arañas
modernas. En estos fósiles se pueden
apreciar, igualmente, tendones musculares.
También se han encontrado fósiles de
arácnidos en la localidad alemana de Alken
an der Mosel, del Devónico inferior (≈400
Ma). Estos fósiles se han conservado de una
forma más típica. Son moldes del animal
vivo, depositados en pizarras formadas en el
lecho de la albufera. Los fósiles de Alken
fueron descritos por Størmer (1970), y entre
ellos está el trigonotárbido Alkenia.
Schultka (1991) describe otro trigonotárbido
de un yacimiento alemán de similar
antiguëdad Rheinischen Schiefergebirges.
Otra localidad importante, del Devónico
medio (≈380 Ma), es Gilboa, en Nueva York.
Gilboa tiene más órdenes de arácnidos
fósiles que ningún otro yacimiento devónico.
Entre sus fósiles hay trigonotárbidos como
Gilboarachne (Fig. 2c), descrito por Shear
et al. (1987). De Gilboa procede el
pseudoescorpión más antiguo que se conoce,
Drachochela (Shear et al., 1989; Schwaller
et al., 1991). También se han encontrado en
Gilboa ácaros, tanto oribátidos (Norton et
al., 1988) como alicorrágidos (Kethley
et
al., 1989). La araña más antigua,
Attercopus, se recogió en Gilboa (Selden et
al., 1991) (P. Selden, en este volumen).
Selden et al. (1991) sugieren que algunos de
los fragmentos que originalmente
describieron como trigonotárbidos pueden
pertenecer a otros órdenes, y así un fósil
incompleto que llamaron Ecchosis podría ser
un amblipígido (araña látigo) primitivo.
Dunlop (en prensa) comenta que algunos
caparazones descritos como trigonotárbidos
se parecen a los de arañas y amblipígidos,
aunque es difícil asociar con seguridad un
caparazón aislado con un orden concreto. Por
otra parte, Selden et al. (1991) sugieren
que puede haber fósiles de arácnidos
primitivos que no pertenezcan a ningún orden
de los actualmente reconocidos, vivo o
extinguido. El "trigonotárbido"
Gelasinatarbus bonamoae, descrito por Shear
et al. (1987) es un ejemplo de tales fósiles
problemáticos.
Se conoce un escorpión del Devónico superior
(≈370 Ma), Hubeiscorpio, de China (Walossek
et al., 1990). No se han descrito
otros arácnidos del Devónico superior,
aunque hay citas de trigonotárbidos de esta
época (W. Shear, comm. pers.) que están a la
espera de ser descritos.

Figura 2:
Arácnidos fósiles del Silúrico y
Devónico. a) Palaeocharinus,
trigonotárbido del Devónico inferior (≈398
Ma) escocés. b) Eotarbus, trigonotárbido
del Silúrico superior (≈414 Ma) de
Shropshire, Inglaterra. c) Gilboarachne,
trigonotárbido del Devónico medio (≈380
Ma) de Gilboa, Nueva York. Las rayas de
escala representan 0'5 mm.
Carbonífero (355-290 Ma)
El Carbonífero se
puede dividir, a grandes rasgos, en
Carbonífero inferior (≈355-325
Ma) y Carbonífero superior (≈325-290 Ma),
equivalentes aproximadamente a los sistemas
americanos Misisípico y Pensilvánico. Como
el Devónico superior, el Carbonífero
inferior presenta muy pocos fósiles de
arácnidos. Se conoce el trigonotárbido
Pocononia, del Carbonífero inferior (≈350
Ma) de Virginia, Estados Unidos (Ewing,
1930; Dunlop, 1996a) y una serie de
trigonotárbidos, aún sin describir, del
Carbonífero inferior de Alemania (R. Rößler,
comm. pers.). Se ha descrito un opiliónido
del Carbonífero inferior (≈335 Ma) de East
Kirkton (Wood et al., 1985). Este es el
opiliónido más antiguo conocido, y aunque
está mal conservado puede apreciarse que
tiene un aspecto moderno y patas largas,
pareciendo un falangioide.
Las primeras investigaciones sobre los
arácnidos del Carbonífero superior se
reflejan sobre todo en las nonografías de
Frie (1904), Pocock (1911) y Petrunkevitch
(1913, 1945a, 1949, 1953, 1955). Una
revisión exhaustiva de la bibliografía
existente sobre los arácnidos del
Carbonífero se saldría de los objetivos del
presente trabajo, por lo que remitimos a
Shear & Kukalová-Peck (1990), Selden (1993a)
y Dunlop (1996c). La mayoría de los fósiles
de arácnidos del Carbonífero superior se
encuentran en asociación con depósitos de
cárbon, y normalmente aparecen en las
pizarras que cubren las vetas de carbón o en
los nódulos redondeados de siderita que se
formaron alrededor del animal cuando murió.
Estos fósiles proceden esencialmente de lo
que parecen haber sido ciénagas. El registro
fósil de arácnidos de este período es el
mejor hasta la llegada de las faunas del
ámbar del Cenozoico.
Los arácnidos del Carbonífero superior
consisten en arañas y escorpiones (descritos
en otros trabajos), y una abundante fauna de
trigonotárbidos. Estos son un grupo
diversificado, en el que se han reconocido
al menos cinco familias y se incluyen
géneros como Anthraromartus,
Anthracosiro, Trigonotarbus, Aphantomartus y
Eophrynus.
Entre ellos hay un ejemplar español (Fig.
3a) de Aphantomartus, descrito por Selden &
Romano (1983) de la provincia de León (≈300
Ma). Dunlop (1996c) propone que el viejo
orden fósil de los Anthracomartida se
considere como sinónimo de los
Trigonotarbida. Además de los Trigonotarbida
hay otros dos órdenes extinguidos (revisados
por Dunlop, 1996c), ambos conocidos sólo del
Carbonífero superior. Los Phalangiotarbida
son un extraño grupo de arácnidos aplanados
(Fig. 4b) con una morfología muy distintiva.
Tienen segmentos opistosómicos reducidos,
piezas bucales muy pequeñas, un tubérculo
ocular con seis ojos y, aparentemente, un
ano dorsal. Beall (1991) da un resumen
reciente de la biología del grupo. Se ha
sugerido que los falangiotárbidos pueden ser
parientes de los opiliónidos (Petrunkevitch,
1949), aunque Dunlop (1995) ha señalado
semejanzas entre los falangiotárbidos y los
ácaros opiliocáridos. El otro orden
extinguido es el de los Haptopoda (Fig. 3b).
Este orden monotípico está representado por
el género Plesiosiro, con sólo nueve
fósiles, todos de un yacimiento inglés. Se
ha sugerido que estos fósiles podrían estar
emparentados con los opiliónidos (Shear &
Kukalová-Peck, 1990), aunque también se
parecen a los uropígidos (Dunlop, 1996c). El
autor prepara en estos momentos una revisión
de Plesiosiro.
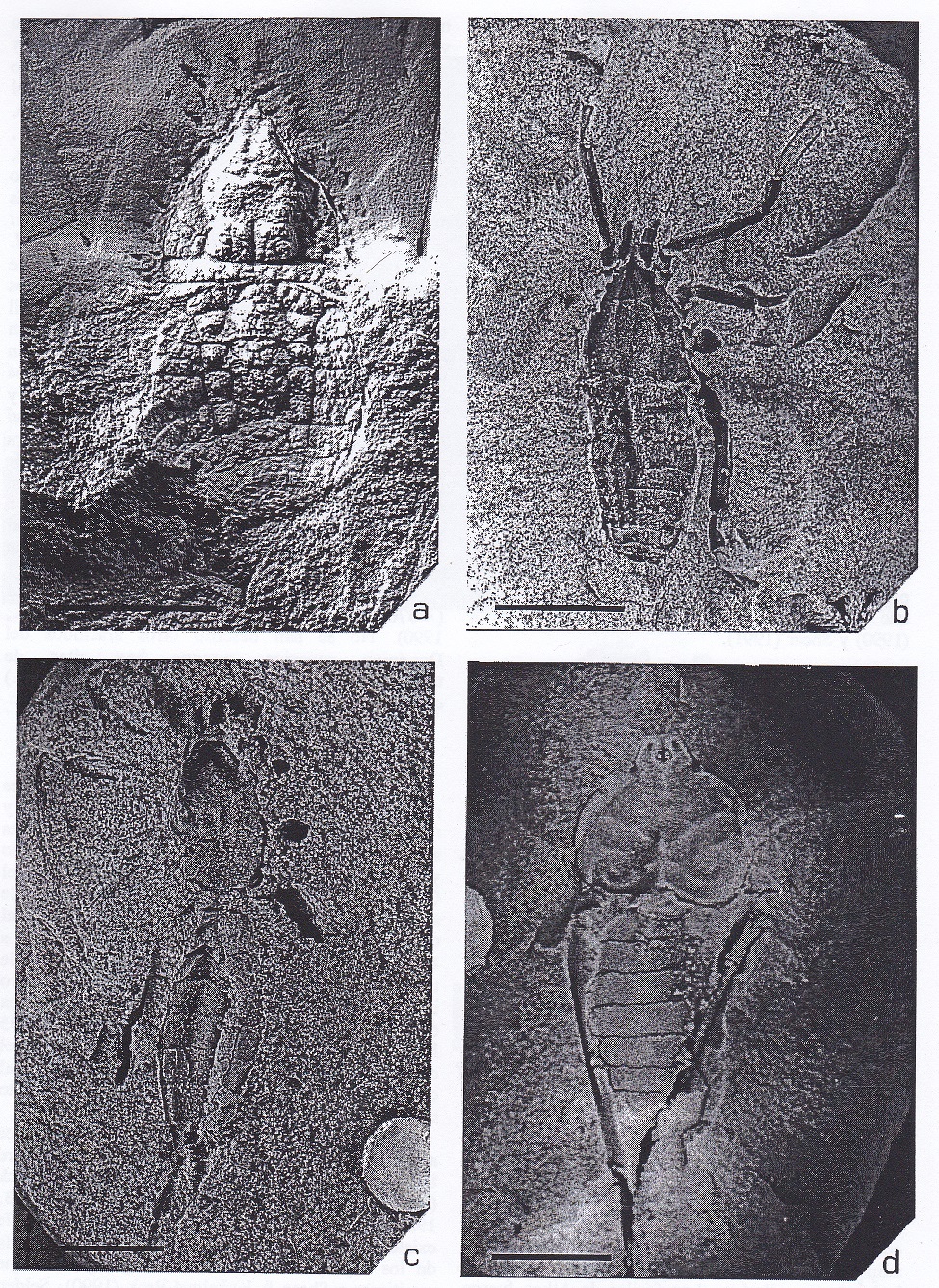
Figura 3:
Arácnidos fósiles del Carbonífero.
a) Aphantomartus. trigonotárbido del
Carbonífero superior (≈380
Ma) de la provincia de León, España. b)
Plesiosiro, haptopódido del Carbonífero
superior (≈305 Ma) de Coseley,
Inglaterra. c) Proschizomus, uropígido
del Carbonífero superior (≈380 Ma) de
Coseley, Inglaterra. d) Graeophonus,
amblipígido del Carbonífero superior
(≈305 Ma) de Coseley, Inglaterra, Las
rayas de escala representan 5 mm.
El Carbonífero superior también ofrece las
primeras apariciones en el registro fósil de
muchos de los órdenes actuales de arácnidos.
Están presentes Uropygi (escorpiones
látigo), como Geralinura, con revisiones
recientes a cargo de Brauckmann & Koch
(1983) y Dunlop & Horrocks (1996), y
Amblypygi (arañas látigo) (Fig. 3d), como
Protophrynus, revisados por Dunlop (1994b).
Los fósiles de estos dos grupos son casi
idénticos a sus representantes actuales.
Dunlop & Horrocks (1996) han identificado un
nuevo género, Proschizomus (Fig. 3c) como
posible grupo hermano de los Schizomida
entre los Uropygi del Carbonífero. Los
Ricinulei del Carbonífero fueron revisados
por Selden (1992). Estos ricinuleidos fueron
divididos en dos grandes bloques, los
Poliocheridae, como Poliocheria, que se
parecen a las formas actuales, con tres
grandes terguitos opistosómicos, y los
Curculioididae, como Curculiodes (Fig. 4a),
que no presentan terguitos visibles y tienen
un surco central a lo largo del opistosoma
que les da el aspecto de escarabajos. Los
Opiliones del Carbonífero (Fig. 4c), como
Nematostomoides, son escasos, y ha habido
pocos estudios sobre ellos desde las
monografías de Petrunkevitch (1953, 1955).
Los fósiles descritos como Kustarachne en un
orden aparte (Kustarachnida) por
Petrunkevitch (1913) son probablemente
opiliónidos (Beall, 1986; Dunlop, 1996c).
Igualmente, algunos de los fósiles del
Carbonífero descritos como arañas pueden ser
también opiliónidos. Hay un único ejemplar
de solífugo del Carbonífero, Protosolpuga,
descrito por Petrunkevitch (1913) de
Norteamérica. El ejemplar fue redescristo
por Selden & Shear (1996), quienes llegan a
la conclusión de que se trata de un
solífugo, aunque en un estado de
conservación demasiado malo como para decir
nada sobre la evolución del grupo.
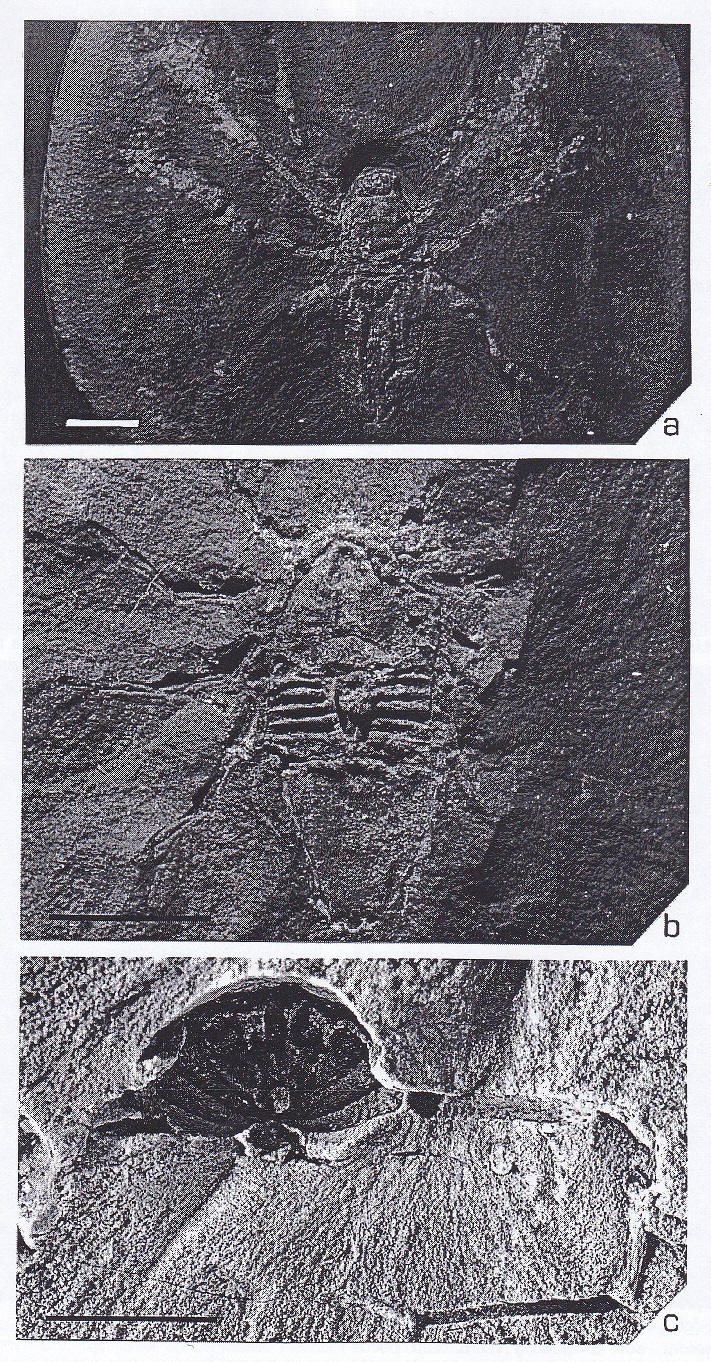
Figura 4:
Arácnidos fósiles del Carbonífero. a)
Curculioides. ricinuleido del
Carbonífero superior (≈305
Ma) de Coalbrookdale, Inglaterra. b)
Mesotarbus, falangiotárbido del
Carbonífero superior (≈305 Ma) de
Westhoughton, Inglaterra. c) Opiliónido
sin determinar del Carbonífero superior
(≈300 Ma) de Montceau-les-Mines,
Francia. Las rayas de escala representan
5 mm.
Pérmico (290-250 Ma)
El material fósil de arácnidos del Pérmico
es muy escaso. Esto puede deberse a una
falta de yacimientos productivos, en
comparación con el Carbonífero. El Pérmico
inferior de Rotliegen, en Alemania (≈280 Ma)
es una excepción, y de él tenemos los
trigonotárbidos más tardíos conocidos
(Müller, 1957). Estos fósiles se han
conservado en pizarras de manera muy
semejante a la de muchos arácnidos del
Carbonífero. También los euriptéridos se
extinguieron durante el Pérmico, y abundan
más los xifosuro de aspecto moderno. Una
hipótesis es que se produce un cambio claro
en las faunas de arácnidos entre el
Paleozoico y el Mesozoico. Los arácnidos
mesozoicos parecen incluir muchos más grupos
de aspecto moderno, especialmente entre las
arañas. No está claro si las arañas llevaron
a los trigonotárbidos a la extinción, o si
cambios ambientales como la desaparición de
ciénagas carboníferas y la expansión de los
reptiles causaron la extinción de los
trigonotárbidos, radiando entonces las
arañas para ocupar los nichos que dejaron
vacíos. Una expansión de los insectos
voladores también habría favorecido la
radiación de las arañas constructoras de
redes.
EL MESOZOICO
Hasta hace algunos años se conocían muy
pocos fósiles de arácnidos del Mesozoico,
con lo que había una gran laguna en el
registro fósil del grupo. Los sistemas
Triásico y Jurásico siguen contando con
pocos fósiles de arácnidos. Los del
Cretácico son ligeramente más abundantes, y
recientemente ha habido más descubrimientos,
por ejemplo en ámbar mesozoico (sobre todo
del Cretáceo) y los fósiles de la formación
Crato, en la provincia de Ceará, en Brasil,
y de Montsec, en España. Los fósiles de
arácnidos mesozoicos consisten
principalmente en arañas (e.g. Selden, 1990;
Selden, en este volumen) y escorpiones del
Triásico (Wills, 1947), aunque hay algunos
representantes, escasos, de otros órdenes de
arácnidos. Los ácaros, especialmente
oribátidos, que son corrientes actualmente
en el suelo (Krivolutsky & Druk, 1986), son
quizás el grupo más común entre los fósiles
del Mesozoico.
Triásico (250-205 Ma)
Se conocen arañas y escorpiones del
Triásico, pero no fósiles de otros órdenes
de arácnidos.
Jurásico (205-135 Ma)
Krivolutsky & Krasilov (1977) citan cinco
especies de ácaros oribátidos, tres
descritas en nuevos géneros (extinguidos),
del Jurásico superior (≈140 Ma) del Este de
la antigua Unión Soviética. Estos autores
sugieren que en esta fauna estaban presentes
oribátidos primitivos y especializados, y
que los oribátidos deben haber radiado antes
del Jurásico. Se ha citado otro ácaro
oribátido del Jurásico inferior (≈201 Ma) de
Suecia (Sivhed & Wallwork, 1978). Estos
autores consiguieron encuadrarlo en un
género actual, Hydrozetes, género que ahora
vive en agua dulce, aunque el fósil se
encontró en un ambiente marino y puede haber
llegado a él por arrastre. Los fósiles de
ácaros oribátidos los han revisado
Krivolutsky & Druk (1986). Un ejemplar de
artrópodo del Jurásico procedente de la
famosa cantera de Solnhofen, en Alemania,
denominado Sternarthron y descrito como
palpígrado por Hasse (1890) ha resultado ser
un insecto y no un arácnido.
Cretácico (135-65 Ma)
Uno de los fósiles de arácnidos más notables
del Cretácico, descrito recientemente, es
Cratosolpuga, un solífugo descrito por
Selden & Shear (1996) del Cretácico inferior
(≈114 Ma) de la formación Crato, en Brasil.
Este fósil, en un excelente estado de
conservación, se podría encuadrar en la
familia actual de los Ceromidae. También hay
ejemplares de un uropígido y un opiliónido,
aún inéditos, de la formación Crato (A.
Jeram, comm. pers.). Se conoce un
pseudoescorpión quernétido del ámbar
cretácico canadiense (Schwaller, 1991), y
del Cretácico de Australia conocemos un
opiliónido de patas largas, con aspecto
moderno (Jell & Duncan, 1986). Se conocen
ácaros oribátidos del Cretácico superior de
Taimyr, Rusia (Bulanova-Zachvatinka, 1974) y
del ámbar cretácico canadiense (McAlpine &
Martin, 1963).

Figura 5:
Cratosolpuga, solífugo del Cretácico
inferior (≈110
Ma) de la Formación Crato, en Brasil.
Las rayas de escala representan 5 mm.
Fotografía cortesía del Dr. Paul Selden.
EL CENOZOICO
El Cenozoico es conocido sobre todo por sus
arañas en ámbar, especialmente las faunas
del ámbar de la República Dominicana y del
Báltico. Hay otras fuentes de ámbar
cenozoico. Hay además material en ámbar de
la mayoría de los otros órdenes de
arácnidos, incluidos escorpiones,
pseudoescorpiones, ácaros, opiliones,
ambipígidos y esquizómidos. Aparte del ámbar
cenozoico hay una serie de fósiles que se
han conservado en pizarras, en condiciones
más típicas. La mayoría de ellos son arañas,
aunque los ácaros oribátidos son también
relativamente comunes, especialmente en
sedimentos más recientes, en los que suele
encontrar el animal entero. Estos ácaros no
son verdaderos fósiles, y a menudo se les
llama subfósiles. La mayor parte de los
arácnidos fósiles del Cenozoico se parecen a
representantes actuales del grupo, y pueden
encuadrarse en familias y/o géneros
actuales. Los fósiles cenozoicos nos dicen
poco sobre la evolución de los taxones
superiores, pero a menudo representan los
registros más antiguos de familias o géneros
actuales de los arácnidos.
Paleógeno (65-23 Ma)
La mayoría de los fósiles paleógenos de
arácnidos se han conservado en ámbar,
especialmente ámbar báltico y dominicano.
Aunque las arañas son sin duda los arácnidos
más corrientes en ámbar; se conoce una
amplia fauna de pseudoescorpiones, y
Schwaller (1982) enumera diez familias para
el conjunto del ámbar cenozoico. Existe un
ejemplo de foresia en un pseudoescorpión
encontrado en ámbar (A. Ross, comm. pers.)
en el que pseudoescorpión se aferra a la
pata de una mosca que se ha posado en el
ámbar. También son corrientes los ácaros
oribátidos en el ámbar báltico (e.g. Koch &
Berendt, 1854; Sellnick, 1918), y se han
descrito igualmente del ámbar mejicano
(Woolley, 1971) y siciliano (Pampaloni,
1902) (ambos ≈25
Ma). Norton & Poinar (1993) re-evalúan los
ácaros del ámbar mejicano y dominicano.
Los opiliónidos son muy corrientes en ámbar,
y en el ámbar báltico se han encontrado unas
15 especies (Scudder, 1891; Roewer, 1939;
Petrunkevitch, 1955; Starega, 1976). También
han sido citados del ámbar dominicano
(Schlee & Glockne, 1978); Cokendolpher,
1986). La fauna de Florissant, Colorado,
Estados Unidos (esta vez no encontrada en
ámbar), fue descrita por Cokendolpher &
Cokendolpher (1982), quienes interpretan
todos estos fósiles como palpatores. Poinar
& Santiago-Blay (1989) describen un
solífugo, Happlodontus, del ámbar
dominicano. Un género actual de
amblipígidos, Tarantula, se conoce también
del ámbar dominicano (Schwaller, 1979). El
esquizómido más antiguo que se conoce se ha
descrito del Oligoceno (≈30 Ma) de China
(Lin et al., 1988), y hay un esquizómido,
aún sin describir, encontrado en el ámbar
dominicano (A. Ross, comm. pers.).
Neógeno (23-1'6 Ma)
Entre los arácnidos fósiles del Neógeno se
encuentran el esquizómido Calcitro
(Petrunkevitch, 1945; Pierce, 1950, 1951) y
Paleokoenia, el palpígrado más antiguo
conocido (Rowland & Sissom 1980). Ambos
provienen del mármol de Onyx, Arizona,
Estados Unidos. Estos dos grupos puede que
hayan existido mucho antes, y probablemente
se remontan al Carbonífero, pero la débil
esclerotización de sus cuerpos hace difícil
su conservación fósil. También se han
descrito ácaros oribátidos de este período
(Krivolutsky & Druk, 1986).
Cuaternario (1'6-0 Ma)
Se han descrito gran cantidad de ácaros
oribátidos de yacimientos cuarternarios
(revisados por Krivolutsky & Druk, 1986), y
estos arácnidos parecen ser relativamente
corrientes en prospecciones y hasta en
restos arqueológicos. Se han encontrado en
sedimentos glaciares (e.g. Nordenskiöld,
1901). A menudo aparecen ácaros subfósiles
en turberas, incluso asociados con los
lugares en que se han encontrado mamuts
(e.g. Coop et al., 1961), tanto en Europa
como en Norteamérica. Los ácaros descritos
tienen normalmente sólo algunos miles de
años de antigüedad, y la mayoría de ellos
pertenecen a géneros y/o especies actuales.
Se podría esperar encontrar
pseudoescorpiones conservados en
circunstancias similares a las de estos
ácaros cuaternarios, pero hasta ahora no se
han localizado. Es posible que hayan
aparecido pero hayan sido pasados por alto.
Entre otros fósiles cuaternarios de
arácnidos tenemos un opiliónido de Italia
(Mastororill, 1965), escorpiones bútidos y
garrapatas ixódidas de Utah y Arizona (Elias
et al., 1992) y ácaros de cuevas de Nuevas
Méjico (Blaszak et al., 1995). Es más
difícil encontrar otros órdenes de arácnidos
de distribución más tropical en el
Cuaternario de Europa, que estuvo dominado
por las glaciaciones.
Discusión
El registro fósil de arácnidos nos puede
decir una serie de cosas importantes sobre
la historia y evolución del grupo. El
material fósil no puede validar o invalidar
un modelo evolutivo concreto, pero
proporciona datos sobre el momento de
aparición, y por consiguiente de divergencia
entre los órdenes de arácnidos,
ofreciéndonos una serie de grupos
extinguidos y fósiles problemáticos que no
parecen encajar en ningún orden conocido,
por lo que pueden resultar cruciales para la
comprensión de la evolución de los
arácnidos. Combinando nuestros conocimientos
sobre el material fósil y las teorías
vigentes sobre la evolución del grupo
(Weygoldt & Paulus, 1979; Shultz, 1990)
podemos proponer las siguientes hipótesis:
1. Los arácnidos
aparecieron, probablemente, a finales del
Cámbrico o principios del Ordovícico.
Parecen tener sus parientes más cercanos en
los Eurypterida (escorpiones de mar), aunque
no tenemos fósiles de arácnidos del
Ordovícico y por tanto no sabemos cómo eran
sus primeros representantes (Dunlop (en
prensa) ofrece una posible recontrucción).
2. Los arácnidos
probablemente llegaron a tierra firme
durante el Silúrico; al menos dos grupos lo
hicieron de forma independiente, los
escorpiones por un lado y el resto de los
arácnidos por otro. Sabemos de escorpiones
acuáticos, pero no sabemos si existían
ejemplos acuáticos de otros órdenes
identificables de arácnidos, o si éstos
evolucionarón una vez que los arácnidos
no-escorpiones llegaron a tierra firme.
3. En el
Devónico ya disponemos de ejemplos
identificables de arañas (Araneae),
escorpiones (Scorpiones), ácaros (Acari),
pseudoescorpiones (Pseudoscorpiones) y del
extinto orden Trigonotárbida. Quizás también
de una araña látigo devónica (Amblypygi), y
de algunos otros fósiles enigmáticos que no
parecen pertenecer a ninguno de los órdenes
actualmente reconocidos.
4. Todos los
órdenes actuales de arácnidos se han
encontrado en el Carbonífero, con la
excepción de los microescorpiones látigo
(Schizomida) y los palpígrados (Palpigradi),
que pueden haber estado presentes, pero no
haber sido hallados aún. Del Carbonífero
conocemos otros dos órdenes extinguidos,
Phalangiotarbida y Haptopoda.
5. Gracias a la
minería de carbón los arácnidos de las
ciénagas carboníferas del período
Carbonífero son los fósiles de arácnidos más
corrientes del Paleozoico. De los fósiles
encontrados, los trigonotárbidos son los más
abundantes, aparte de los escorpiones, y es
posible que fueran uno de los grupos
dominantes de artrópodos predadores
terrestres.
6. La fauna de
arácnidos cambia en la transición del
Carbonífero al Pérmico. Los Haptopoda, los
Phalangiotarbida y luego los Trigonotarbida
desaparecen del registro fósil y
aparentemente se extinguen. Las arañas, y
quizás los opiliones, parecen haber radiado
para ocupar su lugar.
7. El registro
fósil de arácnidos mesosoicos sigue siendo
escaso. Además de arañas y escorpiones, hay
escasos ejemplares de ácaros, opiliones,
solífugos y escorpiones látigo. La mayoría
de ellos son formas de aspecto moderno y nos
iluminan poco sobre la evolución de los
arácnidos durante el Mesozoico.
8. Asimismo, las
fauna cenozoicas, principalmente fósiles
encontrados en ámbar y ácaros subfósiles,
pertenecen en su mayoría a grupos actuales.
Su interés principal reside en evidenciar la
primera aparición de géneros e incluso
especies actuales. La mayoría de las
familias de los arácnidos estaban ya
establecidas, probablemente, para finales
del Mesozoico, aunque hacen falta nuevos
fósiles para confirmar estas hipótesis.
Agradecimientos
Agradezco al Dr
Paul Selden, al Prof. Bill Shear, al Dr
Lyall Anderson, al Dr Simon Braddy, a Andrew
Ross y al Dr Ronny Rößler
sus útiles comentarios. Mi investigación
sobre los orígenes y rediación de los
Chelicerata se ha visto apoyada por una beca
postdoctoral del UK NERC.
Bibliografía
ANDERSON, L.I. y
SELDEN, P.A., (en prensa).-Opisthosomal
fusion and phylogeny of Palaeozoic
Xiphosura. Lethaia.
BEALL, B.S.,
1986.-Reinterpretation of the Kustarachnida
(Abstract). Am. Arachnol., 34: 4.
BEALL, B.S.,
1991.-The Writhlington phalangiotarbids:
their palaeobiological significance. Proc.
Geol. Ass., 102: 161-168.
BERSTRÖM, J.,
1979.-Morphology of fossil arthropods as a
guide to phylogenetic relationships. En:
GUPTA, A.P. (ed.): Arthropod Phylogeny. Van
Nostrand Reinhold Co., New York, pp. 3-56.
BLASZAK, C.,
COKENDOLPHER, J.C. y POLYAK, V.J.,
1995.-Paleozercon cavernicolus, N. gen., n.
sp., fossil mite from a cave in the
southwestern U.S.A. (Acari, Gamasida:
Zerconidae), with a key to Nearctic Genera
of Zerconidae. Inst. J. Acarol., 21:
253-259.
BRAUCKMANN, C. y
KOCH, L., 1983.-Prothelyphonus naufragus n.
sp., ein neuer Gießelskorpion
(Arachnida: Thelyphonida: Thelyphonidae) aus
dem Namurium (unterer Ober-Karbon) von
West-Deutschland. Entomologia Gen., 9:
63-73.
BRIGGS, D.E.G. y
COLLINS, D.A., 1988.-A Middle Cambrian
chelicerate from Mount Stephen, British
Columbia. Palaeontology, 31: 779-798.
BRIGGS, D.E.G., BRUTON,
D.L. y WHITTINGTON, H.B., 1979.-Appendages
of the arthropod Aglaspis spinifer (Upper
Cambrian, Wisconsin) and their significance.
Palaeontology, 22: 167-180.
BRUTON, D.L. y
WHITTINGTON, H.B., 1983.-Emeraldella and
Leancholia, two arthropods from the Burgess
Shale, British Columbia. Phil. Trans. R.
Soc. B, 300: 553-585.
BULANOVA-ZACHVATINKA,
E.M., 1974.-New genera of oribatid mites
from the Upper Cretaceous of Taimyr.
Paleontol. Zh., 2: 141-144 [En ruso].
CASTER, K.E. y BROOKS,
H.K., 1956.-New fossils from the
Canadian-Chazyan (Ordovician) hiatus in
Tennessee. Bull. Am. Paleont., 36: 153-199.
CLARIDGE, M.F. y LYON,
A.G., 1961.-Lung-books in the Devorian
Palaeocharinidae (Arachnida). Nature
(London), 191: 1190-1191.
COKENDOLPHER, J.C.,
1986.-A new species of fossil Pellobunus
from Dominican Republic Amber (Arachnida:
Opiliones: Phalangodidae). Carib. J. Sci.,
22: 205-211.
COKENDOLPHER, J.C. y
COKENDOLPHER, J.E, 1982.-Rexamination of the
Tertiary harvestmen from the Florissant
Formation, Colorado (Arachnida: Opiliones:
Palpatores). J. Paleont., 56: 1213-1217.
COCKERELL, D.T.A.,
1907.-Some fossil arthropods from
Florissant, Colorado. Amer. Mus. Nat. Hist.
Bull., 23:605-616.
COOP, G.P., SHOTTON,
F.W. y STRACHAN, I., 1961.-A late
Pleistocene fauna and flora from Upper
Warren, Worcestershire, Phil. Trans. R. Soc.
Lond., 244: 379-421.
COWIE, J.W. y BASSETT,
M.G., 1989.-International Union of
Geological Sciences 1989 Global
Stratigraphic Chart with Geochronometric and
Magnetostratigraphic calibration. Bureau of
International Commission on Stratigraphy.
DUNLOP, J.A.,
1994a.-Filtration mechanisms in the
mouthparts of tetrapulmonate arachnids
(Trigonotarbida, Araneae, Amblypygi,
Uropygi, Schizomida). Bull. Br. arachnol.
Soc., 9: 267-273.
DUNLOP, J.A., 1994b.-An
Upper Carboniferous amblypygid from the
Writhlington Geological Nature Reverse.
Proc. Geol. Ass., 105: 245-250.
DUNLOP, J.A., 1995.-Are
the fossil phalangiotarbids just big
opilioacarid mites? Newsl. Br. arachnol.
Soc., 74: 8-9.
DUNLOP, J.A., 1996a.-A
redescription of the trigonotarbid arachnid
Pocononia whitei (Ewing 1930). Poläont. Z.,
70: 145-151.
DUNLOP, J.A., 1996b.-A
trigonotarbid arachnid from the Silurian of
Shropshire. Palaeontology, 39: 605-614.
DUNLOP, J.A.,
1996c.-Systematics of the fossil arachnids.
Revue suisse Zool., hors serie: 173-184.
DUNLOP, J.A., (en
prensa).-Palaeozoic arachnids and their
significance for arachnid phylogeny. Proc.
16th Euro. Colloq. Arachnol.
DUNLOP, J.A. y
HORROCKS, C.A., 1996.-A New Upper
Carboniferous whip scorpion (Arachnida:
Uropygi: Thelyphonida) with a revision of
the British Carboniferous Uropygi. Zool.
Anz., 243: 293-306.
DUNLOP, J.A. y SELDEN,
P.A. (en prensa).-The Early History and
Phylogeny of the Chelicerates. En: FORTEY,
R.A. y THOMAS, R.H. (eds.): Proceedings of
the International Symposium on the
Relationships of the Major Arthropod Groups.
Chapman & Hall, Londres.
ELIAS, S.A., MEAD, J.I.
y AGENBROAD, L.D., 1992.-Late Quaternary
arthropods from the Colorado Plateau,
Arizona and Utah. Gt. Basin Nat., 52: 59-67.
EWING, H.E., 1930.-A
fossil arachnid from the Lower Carboniferous
shales (Pocono formation) of Virginia. Ann.
ent. Soc. Am. 23: 641-643.
FLOWER, R.,
1968.-Merostomes from a Cotter horizon of
the El Paso group. Mem. Inst. Min. Tech.,
New Mex., 22(4): 35-44.
FRIC, A.,
1904.-Palaeozoische Arachniden. Publicado de
forma privada por el autor, Praga.
GOULD, S.J.,
1989.-Wonderful Life. Penguin Books,
Londres.
HAASE, E.,
1980.-Breiträge zur Kenntniss der fossilen
Arachniden. Z. dt. geol. Ges., 42: 629-657.
HIRST, S., 1923.-On
some arachnid remains from the Old Red
Sandstone (Rhynie Chert Bed, Aberdeenshire).
Ann. Mag. nat. Hist., 9: 455-474.
HOLM, G., 1898.-Über die organisation des
Eurypterus fischeri Eichw. Mém. Acad. Sci.
St. Petersb., 8(2): 1-57.
JELL, P.A. y DUNCAN, P.M.,
1986.-Invertebrate, mainly insects, from the
freshwater, Lower Cretaceous, Koonwarra
Fossil Bed (Korumburra Group), Soth
Gipplasnd, Victoria. Mem. Ass. Aust.
Palaeo., 3: 111-205.
JERAM, A.J., SELDEN, P.A. y EDWARDS, D.,
1990.-Land animals from the Silurian:
arachnids and myriapods from Shropshire.
Science, 250: 658-661.
KETHLEY, J.B., NORTON, R.A., BONAMO, P.M. y
SHEAR, W.A., 1989.-A terrestrial
alichoragiid mite (Acari: Acriformes) from
the Devonian of New York. Micropaleontology,
35: 367-373.
KOCH, C.L. y BERENDT, G.C., 1854.-Die im
Berstein befindlichen Crustaceen,
Myriapoden, Arachniden und Apteren der
Vorweldt, Publicado por A. MENGE, Berlín,
124 pp.
KRAUS, O., 1976.-Zur phylogenetischen
Stellung und Evolution der Chelicerata. Ent.
Germ., 3:1-12.
KRIVOLUTSKY, D.A. y DRUK, A.Y., 1986.-Fossil
oribatid mites. Ann. Rev. Entomol., 31:
533-545.
KRIVOLUTSKY, D.A. y KRASILOV, B.A.,
1977.-Oribatid mites from Upper Jura
deposits of USSR. En: SKARLATO, O.A. y
BALASHOV, Y.S. (eds.): Morphology and
Diagnostics of Mites. Zoological Institute,
Leningrado, pp. 16-24.
LIN QI-BIN, YAO YI-MIN, XIANG WEI-DA y XIA
YU-RONG, 1988.-An Oligocene
micropalaeoentomofauna from Gubei District
of Shandong and its ecological environment.
Acta Micropal. Sinica, 5: 331-345.
MASTORORILL, V.I., 1965.-I fosilli
Quaternari del Bacino diatomitico di Riano
(Roma) nella collezione del Museo Civico di
Science naturali "D. Doria" in Genova.
Atti
Inst. Geol. Univ. Genova, 3: 1-245.
McALPINE, J.F. y MARTIN, J.E.H.,
1963.-Canadian amber-a palaeontological
treasure chest. Can. Entomol., 101: 819-839.
MÜLLER, A.H., 1957.-Ein Arachnidenrest
(Brachylycosa ? manebachensis n.sp.) aus dem
Unteren Rotliegenden (Manebacher Schichten)
von Thüringen. Geologie, 6: 95-98.
NORDENSKIÖLD, E., 1901.-Zur Kenntnis der
Oribatidenfauna Finlands. Acta Soc. Fauna
Flora Fenn., 21(2): 1-35.
NORTON, R.A. y POINAR, G.O.,
1993.-Reassessment and new records of
oribatid mite fossils from Tertiary
Neotropical amber. carologica, 34: 57-67.
NORTON, R.A., BONAMO, P.M., GRIERSON, J.D. y
SHEAR, W.A., 1988.-Oribatid mite fossils
from a terrestrial Devonian deposit near
Gilboa, New York. J. Paleont., 62: 259-269.
PAMPALONI, L., 1902.-Microflora e microfauna
nel dosodile di Melilli. Rend. Accad.
Lincei. Ser 5, 11: 1-252.
PETRUNKEVITCH, A.I., 1913.-A monograph of
the terrestrial Palaeozoic Arachnida of
North America. Trans. Conn. Acad. Arts Sci.,
18: 1-137.
PETRUNKEVITCH, A.I., 1922.-Tertiary spiders
and opilionids of North America. Trans.
Conn. Acad. Arts Sci., 25: 211-279.
PETRUNKEVITCH, A.I., 1945a.-Palaeozoic
Arachnida of Illinois. An inquiry into their
evolutionary trends. Scient. Pap. Illinois
St. Mus., 3: 1-72.
PETRUNKEVITCH, A.I., 1945b.-Calcitro
fisheri. A new fossil arachnid. Am. J. Sci.,
243: 320-329.
PETRUNKEVITCH, A.I., 1949.-A study a
Palaeozoic Arachnida. Trans. Conn. Acad.
Arts. Sci., 37: 69-315.
PETRUNKEVITCH, A.I., 1953.-Paleozoic and
Mesozoic Arachnida of Europe. Mem. geol.
Soc. Am., 53: 1-122.
PETRUNKEVITCH, A.I., 1955.-Arachnida. En:
MOORE, R.C. (ed.): Treatise on Invertebrate
Paleontology, Part P, Arthropoda 2.
Geological Society of America and University
of Kansas Press, Boulder, Colorado y
Lawrence, Kansas, pp. 42-162.
PIERCE, W.D., 1950.-Fossil arthropods from
the onyx marble. Bull. Sth. Calif. Acad.
Sci., 49: 101-104.
PIERCE, W.D., 1951.-A fossil pedipalpi from
Bonner Quarry. Bull. Sth. Calif. Acad. Sci.,
50: 38-41.
RAASCH, G.O., 1939.-Cambrian Merostomata.
Spec. pap. geol. Soc. Am., nº 19.
ROEWER, C.F., 1939.-Opilioniden im
Bernstein. Palaeobiologica, 7: 1-5.
ROWLAND, J.M. y SISSOM, W.D., 1980.-Report
on a fossil palpigrade from the Tertiary of
Arizona, and a review of the morphology and
systematics of the order (Arachnida:
Palpigradi). J. Arachnol., 8: 69-86.
POCOCK, R.I., 1911.-A monograph of the
terrestrial Carboniferous Arachnida of Great
Bretain. Monographs of the
Palaeontographical Society, pp. 1-84.
POINAR, G.O. y SANTIAGO-BLAY, J.A., 1989.-A
fossil solpugid, Happlodontus proterus, new
genus, new species (Arachnida: Solifugae)
from Dominican amber. Jl. N. Y. ent. Soc.,
97: 125-132.
SCHLEE, D. y GLÖCKNER, W., 1978.-Bernstein.
Stuttgarter Beitr. Naturk., Ser. C, Nº 8, 72
pp.
SCHULTKA, S., 1991.-Trigonotarbus stoermeri
n.sp.- ein Spinnentier aus den Bensberger
Schichten (Ems/Unter-Devon) des Rheinischen
Schiefergebirges. Neues Jb. Geol. Paläont.
Abh., 183: 375-390.
SCHWALLER, W., 1979.-Erstnachweis der
Ordnung Geißelspinnen
in Dominicanischem Berstein (Stuttgarter
Bernsteinsammlung: Arachnida Amblypygi).
Stuttg. Beitr. Naturk. B, 50: 1-12.
SCHWALLER, W.,
1982.-Der erste Pseudoskorpion (Chernetidae)
aus Mexikanischem Bernstein. Stuttg. Beitr.
Naturk. B, 85: 1-19.
SCHWALLER, W.,
1991.-The first Mesozoic pseudoscorpion,
from Cretaceous Canadian amber.
Palaeontology, 34: 971-976.
SCHWALLER, W., SHEAR,
W.A. y BONAMO, P.M., 1991.-The first
Palaeozoic pseudoscorpions (Arachnida,
Pseudoscorpionida). Am. Mus. Novit., 3009:
1-17.
SCUDDER, S.H.,
1891.-Index to known fossil insects of the
world incluiding myriaopods and arachnids.
Rep. U. S. Geol. Surv., 71: 1-744.
SELDEN, P.A.,
1984.-Autoecology of Silurian eurypterids.
Spec. Pap. Palaeont., 32: 39-54.
SELDEN, P.A.,
1990.-Lower Cretaceous spiders from the
Sierra de Montsech, North-East Spain.
Palaeontology, 33: 257-285.
SELDEN, P.A.,
1992.-Revision of the fossil ricinuleids.
Trans. R. Soc. Edinburgh: Earth Sci., 83:
595-643.
SELDEN, P.A.,
1993a.-Fossil arachnids-recent advances and
future prospects. Mem. Qd. Mus., 33:
389-400.
SELDEN, P.A.,
1993b.-Arthropoda (Aglaspidida, Pycnogonida
and Chelicerata). En BENTON, M.J. (ed.): The
Fossil Record 2. Chapman & Hall, Londres,
pp. 297-320.
SELDEN, P.A., y JERAM,
A.J., 1989.-Palaeophysiology of
terrestrialiastion in the Chelicerata.
Trans. R. Soc. Edinburgh: Earth Sci., 80:
303-310.
SELDEN, P.A. y ROMANO,
M., 1983.-First Palaeozoic arachnid from
Iberia: Aphantomartus areolatus Pocock
(basal Stephanian; prov. León, N.W. Spain),
with remarks on aphantomartid taxonomy.
Boln. geol. Min., 94: 106-112.
SELDEN, P.A. y SHEAR,
W.A., 1996.-The first Mesozoic Solifugae
(Arachnida), from the Cretaceous of Brazil
and a redescription of the Palaeozoic
Solifugae. Palaeontology, 39: 583-604.
SELDEN, P.A., SHEAR,
W.A. y BONAMO, P.M., 1991.-A spider and
other arachnids from the Devonian of New
York, and reinterpretations of Devonian
Araneae. Palaeontology, 34: 241-281.
SELLNICK, M., 1918.-Die
Orbatiden der Bernsteinsammlung der
Universität Königsberg. Schr. phys.-ökon.
Ges. Königsb., 59: 21-42.
SHEAR, W.A., 1991.-The
early development of terrestrial ecosystems.
Nature (Londres), 351: 283-289.
SHEAR, W.A. y
KUKALOVA-PECK, J., 1990.-The ecology of
Palaeozoic terrestrial arthropods: the
fossil evidence. Can J. Zool., 68:
1807-1834.
SHEAR, W.A., SCHWALLER,
W. y BONAMO, P.M., 1989.- Record of
Palaeozoic pseudoscorpions. Nature
(Londres), 341: 527-529.
SHEAR, W.A., SELDEN,
P.A., ROLFE, W.D.I., BONAMO, P.M. y
GRIERSON, J.D., 1987.-New terrestrial
arachnids from the Devonian of Gilboa, New
York (Arachnida, Trigonotarbida). Am. Mus.
Novit., 2901: 1-74.
SHULTZ, J.W.,
1990.-Evolutionary morphology and phylogeny
of Arachnida. Cladistics, 6: 1-38.
SIVHED, U. y WALLWORK,
J.A., 1978.-An early Jurassic orbatid mite
from Southern Sweden. Geol. För. Stockh.
Förh., 100: 65-70.
STAREGA, W.,
1976.-Opiliones kosarze (Arachnoidea). Fauna
Polski. Polska Akad. Nauk Instytut Zool.
Warsvawa, 5: 1-197.
STøRMER, L., 1944.-On
the relationships and phylogeny of fossil
and Recent Arachnomorpha. Skr. norske
Vidensk-Akad, nat.-nat. Kl., 5: 1-158.
STøRMER, L.,
1970.-Arthropods from the Lower Devonian
(Lower Emsian) of Alken an der Mosel,
Germany, Part 1, Arachnida. Senckenberg.
leth., 51: 335-369.
STøRMER, L.,
1972.-Arthropods from the Lower Devonian
(Lower Emsian) of Alken an der Mosel,
Germany, Part 2, Xiphosura. Senckenberg.
leth., 53: 1-29.
WALCOTT, C.D.,
1912.-Middle Cambrian Brachiopoda,
Malacostraca, Trilobita and Merostomata.
Cambrian Geology and Palaeontology, II.
Smithson. misc. Collns., 57: 145-288.
WALOSSEK, D., LI, C.-S.
y BRAUCKMANN, C., 1990.-A scorpion from the
Upper Devonian of Hubei Province, China
(Arachnida, Scorpionida). Neues Jb. Geol.
Paläont. Mh., 1990: 169-180.
WEYGOLDT, P. y PAULUS,
H.F., 1979.-Unterzuchungen zur Morphologie,
Taxonomie und Phylogeny der Chelicerata. Z.
Zool. Sys. EvolForsch., 17: 85-116, 177-200.
WILL, L.J., 1947.-A
Monograph of the British Triassic scorpions.
Monographs of the Palaeontolographical
Society, pp. 1-137.
WILL, M.A., BRIGGS,
D.E.G., FORTEY, R.A. y WILKINSON, M.,
1995.-The significance of fossils in
understanding arthropod evolution. Verh. dt.
zool. Ges., 88: 203-215.
WOOLLEY, T.A.,
1971.-Fossil oribatid mites in amber from
Chiapas, Mexico. Univ. Calif. Publ.
Entomol., 63(2): 91-99.
WOOD, S.P., PANCHEN,
A.L. y SMITHSON, T.R., 1985.-A terrestrial
fauna the Scottish Lower Carboniferous.
Nature (Londres), 314: 355-356.
Texto original:
Dunlop, J. A. 1996. Arácnidos Fósiles (con
exclusión de Arañas y Escorpiones) 16,
77-92. ver archivo completo en
PDF
________________________________________________________________________
Complementariamente puedes consultar: