El registro fosil de los Escorpiones:
entre el agua y la tierra
Antonio Melic
1.-Introducción.
Desde antiguo el hombre ha sentido una gran
fascinación por los escorpiones y no ha
dudado en incluirlos en su mitología,
cultura popular y vocabulario (ver Monzón y
Blasco, 1995, 1996a, 1996b, 1996c). Su forma
característica -especialmente el aguijón- y
la peligrosidad de su picadura a
consecuencia del veneno inoculado lo ha
vinculado con fenómenos tanto naturales como
sobrenaturales desde hace al menos 4000
años. Al margen de su importancia cultural o
antropológica y al margen también de su
importancia médica, los escorpiones tienen
un gran valor científico por varios motivos:
(1) Son los quelicerados vivientes más
antiguos que se conocen del registro fósil
y, en consecuencia, constituyen el grupo más
cercano al "arácnido primigénio o ancestral"
por lo que resultan claves para llegar a
entender la evolución y filogenia
tanto de Arachnida como de toda la Clase
Chelicerata y sus parentescos y relaciones
con otros grupos cercanos; (2) Los
escorpiones constituyen uno de los pocos
ejemplos contenidos en el registro fósil en
el que queda documentado el proceso de
terrestrización de formas inicialmente
acuáticas. Este proceso de colonización,
acaecido en el Paleozoico inferior-medio, se
produjo en diversos grupos zoológicos y
grados pero raramente cuenta con secuencias
fósiles en las que aparezcan reflejadas las
sucesivas etapas de adaptación morfológica.
2.-Los Escorpiones actuales.
Scorpionida Latreille 1817, al que
usualmente se le asigna la categoría de
orden, es un grupo de artrópodos
quelicerados (Chelicerata) con el cuerpo
dividido en dos tagmas (ver Fig. 1):
cefalotórax o prosoma y abdomen u
opistosoma, a su vez dividido en dos
regiones: el mesosoma (preabdomen) y el
metasoma (postabdomen). El prosoma está
cubierto por el caparazón (dorsalmente) y
consiste en siete somitos o segmentos
(III-IX), a los que primitivamente debían
unirse dos más (I y II) que en las especies
actuales han desaparecido (y que dan lugar
en otros artrópodos a la cavidad
clipeolabral o preoral y al somito antenal)
(Henry, 1949; Anderson, 1973; Hjelle, 1990).
Los dos primeros segmentos actuales (III y
IV) están representados por los quelíceros y
los pedipalpos respectivamente. Los cuatro
pares de apéndices marchadores o patas
representan a los siguientes cuatro somitos.
El último (IX) sólo presenta apéndices en
estado embrionario (Brauer, 1895;
Abdel-Wahab, 1952; Hjelle, 1990). Tanto los
quelíceros como los pedipalpos están
terminados en pinzas. Los pedipalpos
presentan unas estructuras sensoriales
(tricobotrias) similares a largos y finos
pelos articulados en la base. Ventralmente,
la unión de los apéndices marchadores con el
prosoma forma dos estructuras: las coxas de
los dos primeros pares cierran la cavidad
preoral (o "boca del escorpión") y los dos
últimos rodean al esternón (sternum). La
triangular o transversal) es considerada
como carácter taxonómico en la clasificación
de familias (1).
El preabdomen (mesosoma) es la primera parte
del opistosoma y está compuesto por siete
segmentos el último de los cuales se
estrecha para unirse con el metasoma,
compuesto de otros cinco segmentos y el
telsón. Ventralmente, los dos primeros
segmentos del preabdomen contienen la
obertura genital (X), los peines (XI) y los
estigmas respiratorios o pares de aberturas
que conectan con las filotráqueas (book
lungs) (XII-XVI); el último segmento no
contiene estigmas, apéndices o estructuras
exclusivas de los escorpiones y cuyas
funciones son dudosas. Han sido consideradas
órganos respiratorios, genitales y tactiles
o sensoriales. En general, se ha considerado
que su principal función es mecano-receptora
(captar vibraciones del suelo)
(Cloudsley-Thompson, 1955), pero esta
función parece ser realizada por otros
órganos situados en los basitarsos de las
patas (Brownell, 1985). Se ha señalado
también que pueden ser utilizadas en la
selección de áreas de deposición de los
espermatóforos (Carthy, 1966, 1968) y como
quimioreceptores (Foelix y Müller-Vorholt,
1983). El postabdomen o metasoma consiste en
cinco segmentos (XVII-XXI) más el telsón o
aguijón conectado a glándulas productoras de
veneno y que no es considerado un verdadero
segmento. Estos segmentos portan setas,
pelos y rugosidades de gran valor taxonómico
(Hjelle, 1990).
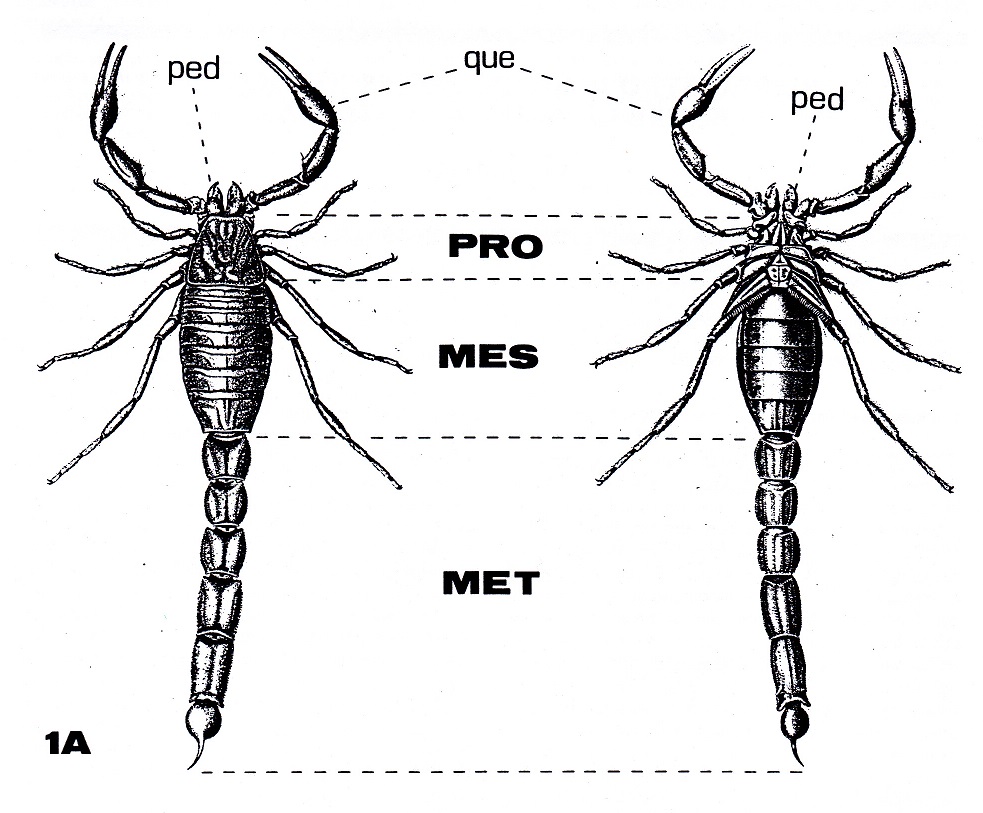
Figura
1A: Morfología externa de un
escorpión moderno (Buthus occitanus).
Leyenda
común a Figura 1A y 1B (pág. siguiente):
cox: coxapoditos. esn:
últimos 5 esternitos del mesosoma.
est: esternum. MES: Mesosoma
o preabdomen. MET: Metasoma o
postabdomen. opg: opérculo
genital. PRO: Prosoma o
cefalotórax. pec: pectinas o
peines. ped: pedipalpos. que:
quelíceros. str: estigmas
respiratorios.
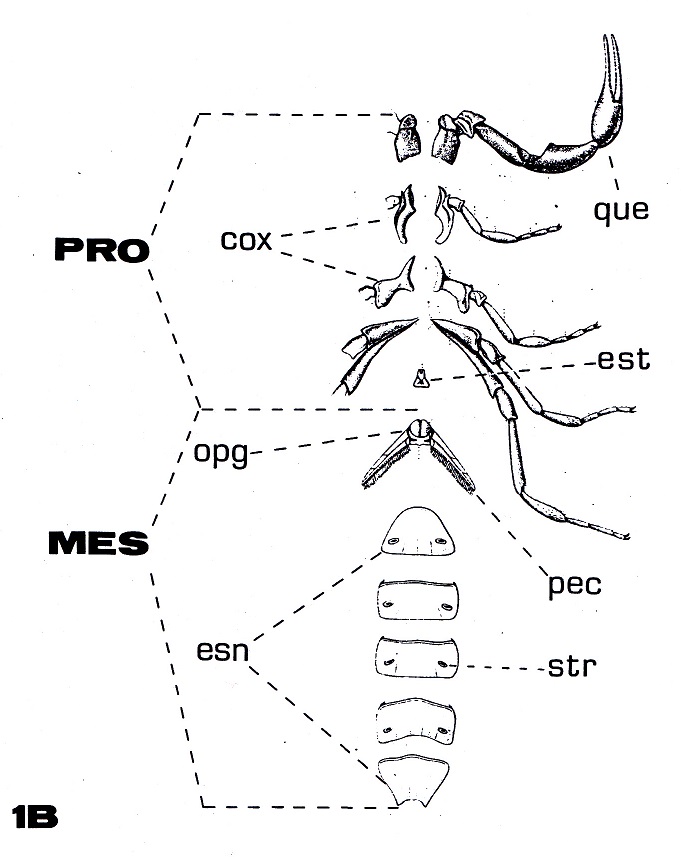
Figura
1B: Prosoma y mesosoma en visíon
ventral de un escorpión actual (ver
leyenda en figura 1A).
Se conocen unas 1400 especies vivientes de
escorpiones pertenecientes a unos 120
géneros englobados en nueve familias
(Sissom, 1990; Stockman, 1991; Dupré, 1995)
que son caracterizadas en base a la
estructura de los quelíceros, tricobotrias,
región coxosternal, espinación de las patas,
caracteres sexuales, etc (ver Sissom, 1990).
Las familias son: Bothriuridae, Buthidae,
Chactidae, Chaerilidae, Diplocentridae,
Ischnuridae, Iuridae, Scorpionidae y
Vaejovidae. El número de superfamilias varía
según los autores. Por ejemplo,
Kjellesvig-Waering (1986) incluye a todos
los escorpiones actuales, junto a otros
extintos, en una sola categoría (el
infraorden Neoscorpionina, ver más
adelante). Birula (1917) y Mello-Leitao
(1945) consideran tres y cuatro
superfamilias respectivamente. Lamoral
(1980) en base a un análisis cladístico de
los caracteres señalados reconoce tres
grandes grupos monofiléticos: Buthoidea
(Buthidae), Chaeriloidea (Chaerilidae) y
Diplocentroidea que incluiría a las
restantes familias, aunque con algunos
problemas pendientes de resolver (ver Fig.
nº 2).
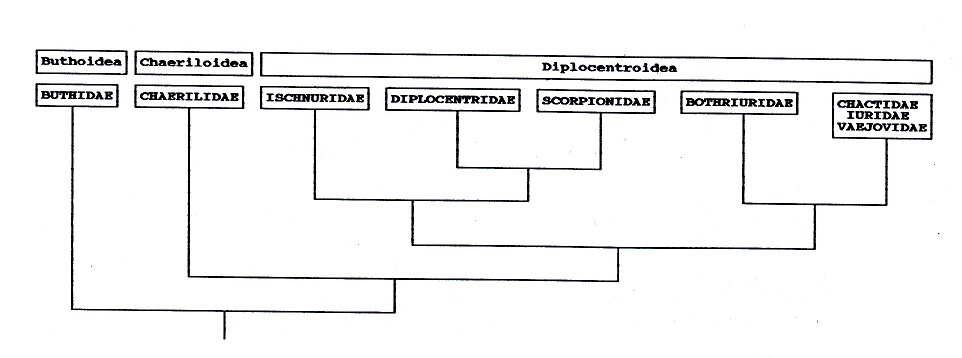
Figura 2:
Hipótesis de la filogenia de las
familias de escorpiones actuales según
Lemoral (1980), modificado por Sissom
(1990); Buthidae sería el grupo hermano
de los restantes escorpiones actuales y
Chaerilidae lo sería de Diplocentroidea.
Este, no obstante, contiene algunas
familias de afinidades inciertas o
problemáticas. Ischnuridae por un lado y
Diplocentridae + Scorpionidae, por otro,
parecen formar un subgrupo monofilético
(Lamoral, 1980; Francke y Soleglad,
1981; Sissom, 1990); en el otro
subgrupo, Bothriuridae es también
monofilético (Lamoral, 1980; Maury,
1980; Francke, 1982), pero la posición
de las restantes tres familias está sin
resolver, "aunque entre ellas pueden
formar otro grupo monofilético (Francke
y Soleglad, 1981.
Respecto a la biología de los escorpiones
puede consultarse Polis (1990); aquí sólo
nos interesa destacar que todas las especies
actuales son terrestres sin excepción y
depredadoras. El interés y problemas
relacionados con la biografía de los
escorpiones han sido recientemente tratados
en Lourenço (1996), donde se aporta
informacion paleobiogeográfica.
3.-El registro fósil de Scorpionida.
Se conocen un centenar de especies fósiles.
A pesar de que esta cifra no resulta
importante, el registro se extiende durante
más de 400 millones de años, del Silúrico
inferior a la actualidad. La Fig. nº 3
resume la cronología de Scorpionida según
los datos disponibles del registro fósil. La
Tabla nº 1 resume a su vez el nivel de
diversidad filogenética alcanzado por el
grupo en cada uno de los períodos
geológicos. Los datos han sido extraídos de
Kjellesvig-Waering (1986), Sisson (1990) y
Selden (1993a), entre otros. Los niveles
taxonómicos, así como la propia
clasificación del grupo, no son precisamente
"pacíficos". Se basan en la obra póstuma del
primero de los autores citados y aunque ha
sido seriamente criticada (por ejemplo por
Stockwell, 1989; Selden, 1993a, 1993b;
Jeram, 1994a, 1994b, entre otros), lo cierto
es que las anteriores (Petrunkevitch, 1949,
1953, 1955) tampoco resultan satisfactorias
(2) (Størmer,
1976; Kjellesvig-Waering, 1986;
Selden, 1993b).
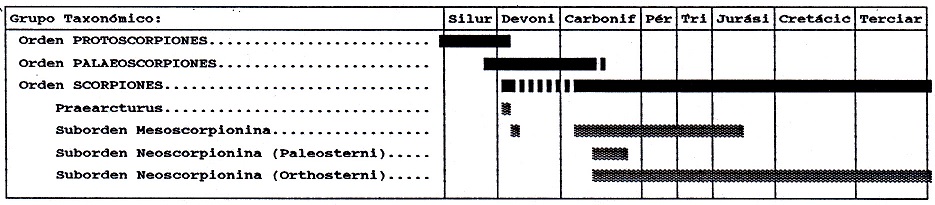
Figura 3:
Cronología de los escorpiones según el
registro fósil. Datos de Selden (1993a).
|
NIVEL
Período Geológico |
|
TAXONOM. |
SILUR. |
DEVON. |
CARBO. |
PERMI. |
TRIAS. |
JURAS. |
CRETA. |
TERCI. |
|
Superfam. |
6 |
6 |
14 |
1 |
4 |
1 |
1 |
2 |
|
Familia |
6 |
7 |
30 |
1 |
5 |
2 |
1 |
2 |
|
Género |
7 |
8 |
45 |
1 |
5 |
2 |
1 |
6 |
Tabla
1: Diversidad supraespecífica en
cada uno de los períodos geológicos
(basado en datos de Sissom, 1990 y
otros)
De la figura y cuatro anteriores pueden
extraerse una serie de consecuencias: (1) La
gran antigüedad de Scorpionida en el
registro fósil; (2) Su participación en el
proceso de colonización del medio terrestre
(acaecido entre el Silúrico y el Devónico);
(3) Su gran diversidad "filogenética"
(taxones supraespecíficos) en el Paleozoico
a pesar del relativamente bajo número de
especies fósiles conocidas, en comparación
con la actual; (4) Su escasa representación
en el Mesozoico, lo que por otro lado
resulta habitual en otros grupos (ver, p.e.,
Arachnida o Myriapoda, en este Volumen).
En las líneas que siguen vamos a intentar
trazas un breve recorrido por la cronología
del grupo a lo largo del Fanerozoico.
Silúrico
El origen de los escorpiones es incierto,
como lo es su filogenia y relaciones con
otros grupos de quelicerados (ver más
adelante). El más antiguo escorpión fósil
conocido es Dolichophonus loudonensis
(Laurie, 1899), del Silúrico inferior de
Escocia. De finales del período son
conocidos representantes de otros géneros de
escorpiones: Proscorpius, Archaeophonus,
Stoermeroscorpio, Brontoscorpio (con formas
gigantes como B. anglicus de 94 cm) o
Palaeophonus, uno de cuyos representantes
(P. nuntius Thorell y Lindström, 1884, de
Suecia) fue "reconstruido" por Pocock (1901)
y reproducimos en la Fig. nº 4.
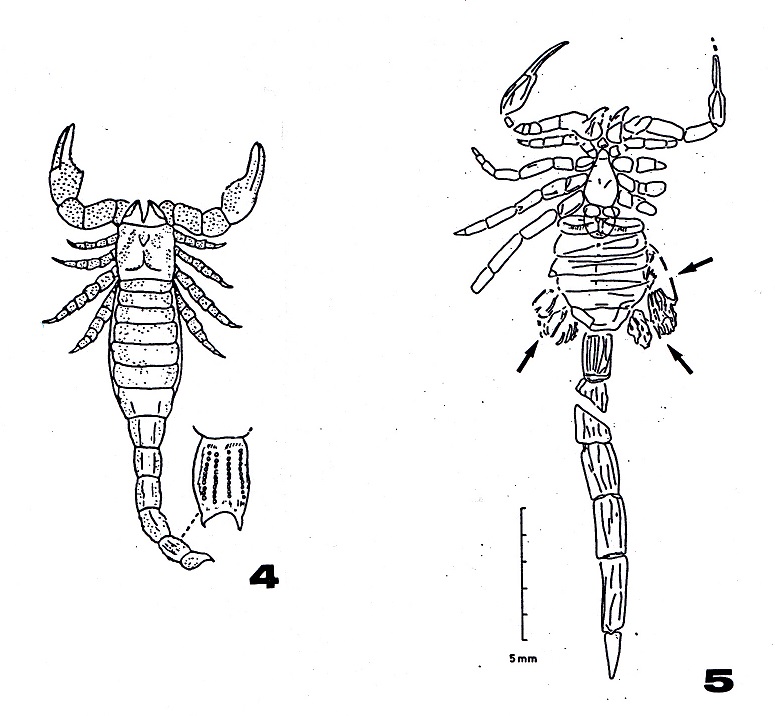
Figura 4:
Reproducción de Palaeophonus nuntius
Thorell y Lindström, 1884, de Pocock
(1901).
Figura 5:
Waeringoscorpio hefteri Størmer, 1970,
del Devónico Inferior (Alemania). Las
estructuras señaladas con flechas
corresponden a las branquias, lo que
indica un modo de respiración acuática.
El escorpión D. loudonensis ha sido
considerado comúnmente como el primer
artrópodo terrestre, a pesar de que Pocock
(1901) ya apuntó la posibilidad de que fuera
una especie acuática al carecer de estigmas
respiratorios. Esta ausencia no constituyó
una prueba para Petrunkevitch (1949, 1953)
ni para Millot y Vachon (1949). Sin embargo,
el descubrimiento posterior de
Waeringoscorpio por Størmer
(1970), del Devónico, y de unas estructuras
situadas en el preabdomen de algunos
ejemplares que resultaron ser branquias (ver
Fig. nº 5) han permitido reinterpretar la
morfología y modo de vida de los escorpiones
del Silúrico (3). Estas branquias se
ubicaban en los cinco primeros segmentos del
preabdomen bajo unas placas abdominales de
protección situadas por encima de los
esternitos. De esta forma, aunque se dispuso
durante años de especímenes fosilizados
visibles ventralmente las branquias
(estructuras filamentosas muy delicadas)
quedaban ocultas en el estuche formado por
la placa y el esternito. En el caso de
Waeringoscorpio las branquias se encontraban
fuera del estuche, lo que permitió su
descubrimiento.
Si nos fijamos de nuevo en la figura 4
comprobaremos que la morfología general de
Palaeophonus nuntius (y lo mismo ocurre con
los restantes géneros) es muy similar a la
de las especies actuales, ninguna de las
cuales es acuática o anfibia. Sin embargo
existen importantes diferencias que delatan
un modo de vida diferente. Por ejemplo la
estructura de las patas marchadoras, cortas
y poco aptas para el desplazamiento
terrestre, así como su terminación en un
tarso cónico (en lugar de las uñas tarsales
de las especies de vida aérea) indican que
estos escorpiones estaban más adaptados al
medio acuático donde el peso del cuerpo al
desplazarse es un problema menor (Pocock,
1911; Størmer, 1976). Otra diferencia
significativa es la presencia de setas
sensoriales de diverso tipo en los
pedipalpos (Wills, 1959, 1960; Størmer,
1963, 1976; Kjellesvig-Waering 1966, 1986)
en lugar de tricobotrias. Estas setas son
pequeñas excrecencias relativamente gruesas
y cortas. En consecuencia, estructuras más
adecuadas para el medio acuático, en el que
los débiles pelos sensores estarían
sometidos a un gran rozamiento y presión y
tendrían muy poca utilidad.
Los escorpiones paleozoicos presentan otras
diferencias morfológicas con los actuales
además de las señaladas que pueden resumirse
en: (1) Una gran variabilidad en la posición
de las apófisis coxales con relación al
esternum, relativamente homogénea en los
escorpiones actuales; (2) En algunos casos,
presencia de ojos compuestos (en las
especies vivientes son simples o están
ausentes); (3) Preabdomen compuesto por 8
segmentos en lugar de 7. Esta característica
se mantiene exclusivamente en los estados
embrionarios de las especies actuales,
aunque luego desaparece (Polis y Sissom,
1990).
Como conclusión, todos
los escorpiones conocidos del Silúrico
parecen ser marinos o habitantes de
sedimentos marinos marginales (lagos
salados, etc.) ya que su morfología sugiere
un modo de vida acuático (Wills, 1947;
Størmer, 1970, 1976; Rolfe y Beckett, 1984;
Kjellesvig-Waering, 1986; Selden, 1993b).
Devónico
El registro fósil del Devónico recoge la
aparición de nuevos géneros de escorpiones:
Branchioscorpio, Hydroscorpius y
Acanthoscorpio de Wyoming (EE. UU.) y
Palaeoscorpius de Alemania. Praearcturus
gigas Woodward, 1871, con un tamaño
aproximado de un metro, tal vez terrestre, y
de posición taxonómica incierta (Selden,
1993a), es descrito de Inglaterra. Størmer
(1970) describe Waeringoscorpio heiteri de
Alken an der Mosel (Alemania) al que ya nos
hemos referido anteriormente
(4),
en un yacimiento en el que la fauna
dominante son euriptéridos (también
conocidos como "escorpiones marinos", ver
Dunlop, 1996, en este volumen). Entre ambos
grupos de Chelicerata existen grandes
semejanzas que permiten presumir una cierta
proximidad filogenética (ver, p.e. Rolfe,
1985).
Varios autores han manajado la hipótesis de
que la terrestrización de varios grupos de
animales se produjo en el Devónico o a
finales del Silúrico
(5) en oleadas sucesivas. Respecto a
los escorpiones, Selden y Jeram (1989),
sugieren que las primeras especies
"pulmonadas" surgen a finales del Devónico a
partir de formas anfibias presentes desde
finales del Devónico a partir de formas
anfibias presentes desde finales del
Silúrico (Shear y Kukalová-Peck, 1990). En
general resulta difícil precisar el modo de
vida de una especie fósil y la variedad de
diseños de órganos respiratorios y su
difícil conservación hacen complicado
determinar el momento preciso de transición
entre formas acuáticas y terrestres (Størmer,
1976; Shelden y Jaram, 1989; Selden, 1993b).
Así, un análisis de las estructuras
respiratorias de los euriptéridos (Selden,
1985) sugiere que algunas especies eran
capaces de sobrevivir temporalmente en
tierra firme como anfibios. Entre ambos
modos de vida, es razonable pensar que los
antiguos escorpiones (al menos algunas
especies) fueron terrestrizándose
progresivamente, adaptándose a mayores
tiempos fuera del agua (Robinson, 1987), lo
cual pudo conseguirse gracias a las placas
abdominales protectoras que, como ocurre con
algunos xifosuros actuales, les permitían
mantener húmedas las branquias durante las
excursiones terrestres
(6). De los escorpiones citados, sólo
Palaeoscorpius ha sido localizado en
sedimentos marinos profundos;
Waeringoscorpio vivía en un lago hipersalino
conectado con el mar sólo en períodos de
marea alta; los restantes han sido hallados
en sedimentos de estuarios o depósitos
fluviales (Rolfe, 1985). Praearcturus
presenta además una rara peculiaridad: las
coxas de sus pedipalpos disponen de un
órgano de estridulación que parece ser una
adaptación al medio terrestre (Rolfe, 1985).
La prueba definitiva de un modo de vida
terrestre es, evidentemente, la posesión de
órganos de respiración aérea que, en el caso
de los escorpiones, toma forma de
filotráqueas (o pulmones en libro = book
lungs). Estas no aparecen con seguridad
hasta el Carbonífero. Sin embargo, el
análisis de restos fragmentarios de la
cutícula y otras estructuras de los
yacimientos canadienses (Shear et al., 1996)
ha facilitado la evidencia de la presencia
de filotráqueas en escorpiones del Devónico
inferior (Emsiense).
Carbonífero
Este período constituye la "edad de oro" de
los escorpiones, pues alcanzan el mayor
grado conocido de diversidad
supraespecífica. Con arreglo a la
clasificación de Kjellesvig-Waering (1986)
el registro fósil detecta la presencia de 30
familias pertenecientes a 14 superfamilias.
El número de géneros se acerca a 50. Entre
los géneros presentes pueden citarse
Archaeoctonus y Pseudoarchaeoctonus de
Escocia (primeros representantes de una
superfamilia que incorpora lóbulos maxilares
en el primer par de coxas y tal vez
anfibios), Phoxiscorpio (el primer
mesoscorpión en la clasificación de
Stockwell [1989] (7),
de Escocia, con dos pares de coxas por
delante del esternón, aunque sólo el primero
lobulado y vida terrestre), Gigantoscorpio
(con formas gigantes, como su nombre
indica), Isobuthus (de Checoslovaquia),
Eoscorpius (de Inglaterra y EE.UU.) o
Titanoscorpio (de afinidad incierta). Otros
escorpiones terrestres exclusivos del
Carbonífero son Allobuthiscorpius
(Inglaterra), Buthiscorpius (de Francia) y
otros géneros que conforman la superfamilia
Eoctonoidea que, para otros autores
(Stockwell, 1989) tiene el valor de
Infraorden (Palaeosterni) por presentar
algunas características "modernas": grupos
de ojos laterales reducidos y dos pares de
coxas modificados (con lóbulos maxilares)
por delante del esternón.
Jeram (1990b) señala la presencia de
filotráqueas -y en consecuencia, de
respiración aérea y vida terrestre-. en el
Carbonífero en base a dos especímenes
fósiles de Escocia. En las especies actuales
las filotráqueas se conectan con el exterior
a través de aberturas situadas en los
esternitos del preabdomen. Estas, conocidas
como estigmas respiratorios, se presentan
por pares en sólo cuatro esternitos a
diferencia de las placas abdominales
protectoras de las branquias en las especies
acuáticas, que lo hacen en número de cinco.
Kjellesvig-Waering (1986) dividió el orden
Scorpionida en dos subórdenes en función de
la presencia o no de dichas placas.
Branchioscorpionina incluiría a todas
aquellas especies con placas abdominales y
branquias. Neoscorpionina incluiría a las
especies terrestres, con filotráqueas y sin
placas ( y en consecuencia, a todos los
escorpiones actuales). Sin embargo, como ya
hemos indicado, la clasificación de
Kjellesvig-Waering es confictiva y no es
aceptada por todos los paleontólogos. Uno de
los motivos es la distinción anterior. Jeram
(1990b, 1994a) al igual que Shear et al.
(1996) señalan la presencia tanto de placas
como de filotráqueas en las mismas especies,
faltando, en el primer caso, los estigmas
respiratorios. Estos no aparecen con
seguridad hasta Palaeopisthacanthus
schuchertii Petrunkevitch, 1913, el único
escorpión paleozoico con estas aberturas
(Vogel y Durden, 1966; Jeram, 1994b). Otro
género interesante, también perteneciente a
Neoscorpionina, es Compsoscorpius elegans
Petrunkevitch, 1949 por presentar ojos
laterales reducidos.
Estudios sobre la morfología y sistemática
de los escorpiones del Carbonífero pueden
consultarse en Wills (1959, 1960) y Jeram
(1990, 1994a, 199b).
Pérmico + Mesozoico
El registro fósil de escorpiones se reduce
sustancialmente desde finales del Paleozoico
hasta el Terciario. Entre los escorpiones
Branchioscorpionina pueden citarse:
Feistmantelia (Pérmico, Checoslovaquia),
Stenoscorpio, Willsiscorpio, Spongiophonus y
Bromsgroviscorpio (Triásico, Inglaterra),
Mesophonus (Triásico-Jurásico, Europa) y
Liassoscorpionides (Jurásico, Alemania). Del
suborden Neoscorpionina, sólo se conoce
Araripescorpio ligabuei Campos, 1986, de la
Formación Santana (Brasil), incluido en la
Superfamilia Scorpionoidea (ver Selden,
1993a), único escorpión fósil del Cretácico.
Los escorpiones del Triásico inglés han sido
estudiados por Wills (1947). Una de las
especies del período (Mesophonus gracilis
Wills) presenta unas aberturas elípticas en
la parte posterior de las placas lo que
sugirió tanto a Wills como a Størmer
(1963) la posibilidad de que éstas fueran
especies de transición entre los escorpiones
del Paleozoico y las formas modernas, aunque
otros autores consideran que se trata de un
caso de convergencia (Stockwell, 1989;
Jeram, 1994b). Junto a otros géneros como
Phoxiscorpio, Mazonia, Isobuthus o Eobuthus
conforman en la clasificación de Stockwell
(1989) el suborden Mesoscorpionina
(mesoscorpiones), un conjunto de especies
que presentan características más
evolucionadas que Protoscorpionida y
Palaeoscorpionida (filotráqueas, número de
placas abdominales inferior a cinco, cavidad
preoral presente y otras novedades
relacionadas con la terrestrización de su
modo de vida), pero todavía lejos de las
formas actuales (que presentan estigmas
respiratorios, tricobotria muy desarrollada,
reducción ocular...).
Terciario
Millot y Vachon (1949) ya señalan que
resulta difícil separar las especies
actuales de las fósiles del Terciario. Y
efectivamente, todas han sido asignadas a
Buthoidea (Bu) y Scorpionoidea (Sc). Hasta
hace muy pocos años sólo tres especies eran
conocidas del Terciario: (1) Del ámbar
Báltico del Oligoceno (≈35
m.a.) dos ejemplares juveniles de Tityus
eogenus Menge, 1854 (Bu); (2) Del mismo
ámbar y época se conoce Scorpio schweiggeri
Holl, 1829 (Sc?/Bu?); (3) Del Mioceno (≈25
m.a.) de Alemania Mioscorpio zeuneri Hadzi,
1931 (Sc). Recientemente han sido descritos
algunos nuevos escorpiones. En concreto del
ámbar dominicano Centruroides beynai
Schawaller, 1979 (en base a dos juveniles),
Microtityus ambarensis (Schawaller, 1982),
inicialmente descrito en el género Tityus
(ver Santiago-Blay et al., 1990) y Tityus
geratus Santiago-Blay y Poinar, 1988. Todos
ellos pertenecientes a Buthoidea. Otro
Centruroides ha sido descrito, aunque con
dudas, del ámbar mejicano (Santiago-Blay y
Poinar, 1993) (8).
También del ámbar Báltico se conoce
Paläolychus balticus (ver Ganzelewsky y
Slotta, 1996). Por último, del Mioceno se
conoce también Sinoscorpio shanwangensis
Hong, 1983 de la Formación Shanwang,
Shandong Province (China) (Hong, 1983, 1985).
4.-Posición de Scorpionida entre los
quelicerados.
Los escorpiones ocupan un importante lugar
en la interpretación de Chelicerata debido a
su antigüedad (Sissom, 1990).
Tradicionalmente se ha considerado que
Chelicerata incluye tres grupos: (1)
Merostomata (Xiphosura + Eurypterida, es
decir, las especies acuáticas), (2)
Arachnida (escorpiones y otros arácnidos) y
(3) Pantopoda (=Pycnogonida o arañas de
mar). La afinidad del último grupo es
oscura. Respecto a Merostomata, parece
tratarse de un grupo parafilético y sin
valor taxonómico (ver Dunlop, 1996, en este
volumen). Selden (1990, 1993b) ha comentado
tres diferentes hipótesis filogenéticas de
Chelicerata resultantes de otros tantos
análisis cladísticos (Weygoldt y Paulus,
1979; Hammen, 1989 y Shultz, 1990). Según la
hipótesis propuesta por Weygoldt y Paulus
(1979) y compartida por Boudreaux (1979)
(ver Fig. nº 6), tanto Xiphosura primero
como Eurypterida posteriormente se separan
del resto de Chelicerata en un período muy
temprano. Eurypterida constituye el grupo
hermano de Arachnida (Escorpiones + Otros
arácnidos). Este esquema sitúa a Eurypterida
muy cerca de Scorpionida. Existen
importantes caracteres que indican una gran
proximidad filogenética entre ambos grupos,
aunque algunos de ellos pueden ser meramente
convergentes (9)
(Størmer, 1963;
Boudreaux, 1979; Weygoldt y Paulus, 1979;
Kjellesvig-Waering, 1986): segmentación
corporal y apéndices, reducción del prosoma,
posesión de placas abdominales protectoras
de branquias, metasoma de cinco segmentos
terminado en telsón, macrosetas sensoriales,
ojos laterales compuestos, etc. Las
homologías, ojos laterales compuestos, etc.
Las homologías o sinamopomorfías se
producen, en realidad, con los escorpiones
más antiguos del Paleozoico
(Brachioscorpionina). Por contra sólo
existen tres autopomorfías que separan ambos
grupos: chelas de los pedipalpos, y
glándulas productoras de veneno y peines o
pectinas. Las glándulas venenosas no son un
carácter definitivo, pues resulta difícil
precisar si Eurypterida las poseía o no. Las
pectinas, sin embargo, son estructuras
únicas de Scorpionida, aunque algunos
autores (Savory, 1977) las consideran
derivadas de las branquias lamellosas de
Xiphosura. Su función, como ya hemos
comentado anteriormente, es enigmática,
especialmente si consideramos que ya existe
en las formas más antiguas marinas, por lo
que su función actual -en el medio
terrestre- puede ser incluso diferente de la
inicial.
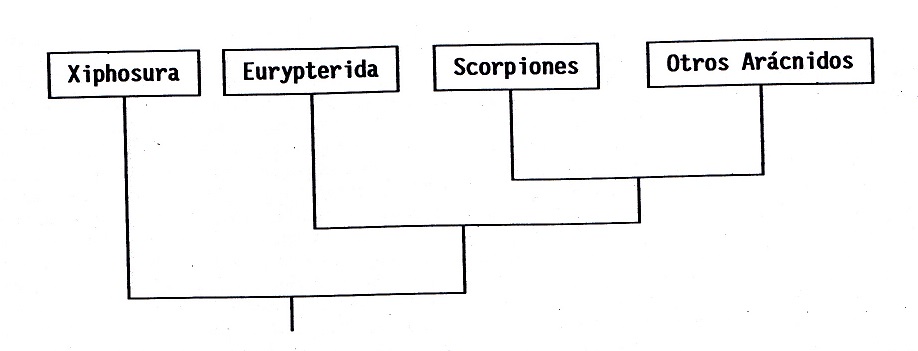
Figura 6:
Filogenia simplificada de Chelicerata
según Weygoldt y Paulus (1979). Esta
hipótesis, incluso en lo relativo a la
mayoría de los "otros arácnidos", es
congruente con la cronología del
registro fósil.
La hipótesis filogenética de Weygoldt y
Paulus (1979) tiene la virtud de ser
congruente con la cronología del registro
fósil (Selden, 1990): Xiphosura aparece en
el Cámbrico (≈550 m.a.), Eurypterida en el
Ordovícico (≈500 m.a.), Scorpionida en el
Silúrico (≈450 m.a.) y los restantes
arácnidos en el Devónico (≈400 m.a.).
Los escorpiones parecen ser el grupo hermano
de los restantes Arachnida según Weygoldt y
Paulus (1979), pero esta relación no está
exenta de problemas. Los autores señalan
cuatro caracteres primitivos (sinapomorfías)
que los relacionan (filotráqueas, tubos de
Malpighi, cavidad preoral para digestión
extraintestinal y pérdida de los ojos
compuestos); sin embargo, este análisis
considera a los escorpiones como animales
terrestres, con lo que la coincidencia sólo
podría establecerse con las familias
"modernas" (Neoscorpionina y algunas otras
familias paleozoicas). El carácter derivado
de Scorpionida sería la posesión de peines o
pectinas. Otros autores consideran la
existencia de un ancestro diferente para
ambos grupos de Arachnida (Anderson, 1973;
Bergström, 1979; Hammen, 1989), lo que, a su
vez, implicaría que la colonización
terrestre de los arácnidos se produjo en
varias ocasiones y momentos diferentes.
5.-Tendencias evolutivas en Scorpionida
Kjellesvig-Waering (1986) lista una serie de
tendencias en la evolución del grupo que, en
gran medida, están directamente relacionadas
con la transición del modo de vida acuático
a terrestre. Estas tendencias, también
señaladas por Størmer (1976), puede
resumirse en: (1) Cambio en las estructuras
y órganos respiratorios, lo que lleva a la
pérdida de las branquias que son
"sustituidas" por filotráqueas. Las placas
abdominales protectoras de las branquias
pierden su utilidad y tienden a desaparecer,
lo que provoca la desaparición de uno de los
esternitos del mesosoma (de ahí que
Petrunkevitch separara a los escorpiones en
dos grandes grupos según poseyeran 8 o 7
esternitos). Así, mientras que las especies
antiguas tienen cinco placas abdominales,
las modernas sólo poseen cuatro esternitos
con filotráqueas. Este proceso trae consigo
la aparición de aberturas de comunicación
con el exterior que se produce a través de
los estigmas respiratorios característicos
de las especies más modernas (incluidas las
actuales) que atraviesan el propio
esternito. (2) Modificaciones en la parte
ventral del prosoma donde donde se unen las
coxas y el esternum. Las formas más arcaicas
presentan un esternum relativamente alargado
que separa las dos zonas donde se unen las
coxas al prosoma. Las más recientes tienden
a cambiar esta morfología dejando el
esternum encerrado exclusivamente entre los
dos últimos pares de coxas; los dos primeros
pares quedan por delante del conjunto y
tienden a desarrollar lóbulos (coxapoditos o
apófisis coxales) que intervienen en la
formación de una cavidad preoral o tubo
relacionado con la alimentación y digestión
de los escorpiones. Ello es básicamente una
consecuencia del proceso de adaptación al
medio terrestre relacionado con las
diferentes condiciones físicas en que debe
producirse la alimentación en uno y otro
medio. Relacionado también con las
necesidades alimentarias, se produce una
reducción de los quelíceros. (3)
Modificaciones en los órganos sensoriales.
Entre ellas se cuentan la transformación de
macrosetas (en especies acuáticas/anfibias)
en tricobotrias, que tienden a aumentar un
número en las especies modernas. Estos
cambios son entendidos por los autores como
signos de adaptación al medio terrestre. (4)
Desaparición de los ojos laterales
compuestos y su sustitución por grupos de
ojos simples que, además, tienden a
reducirse en su número, así como el
desplazamiento hacia atrás de los ojos
medios, lo que se considera una adaptación a
la forma de vida nocturna (habitual en las
especies modernas) frente a lo que debió ser
una generalizada actividad diurna en las
especies del Carbonífero (Jeram, 1994a).
Esta disminución del potencial visual de los
escorpiones se vería, a su vez, compensada
con un aumento de otras estructuras
mecanoreceptoras y/o quimioreceptoras (p.e.,
tricobotrias). (5) Existen otras tendencias
que afectan a los apéndices involucrados en
el movimiento (reducción en tarsos), pérdida
de espinas, etc., relacionadas directamente
con la terrestrización.
6. Clasificación y filogenia de
Scorpionida (incluyendo especies extintas).
Los escorpiones parecen ser un grupo
monofilético, pero presentan algunos
problemas que impiden descartar la hipótesis
de la polifilia. Petrunkevitch (1953)
estableció dos subórdenes (Protoscorpionina
y Euscorpionina) en base al número de
segmentos abdominales (preabdomen de 8 o 7
terguitos respectivamente).
Kjellesvig-Waering, por su parte, estableció
dos subórdenes: Branchioscorpionina que
comprendería a todos los escorpiones con
branquias, es decir, a las especies que
obtienen el oxígeno del agua, y
Neoscorpionina que incluiría a las especies
terrestres (con respiración aérea a través
de filotráqueas). El primer suborden se
divide a su vez en cuatro infraórdenes en
función de la estructura de las placas
abdominales protectoras de las branquias; el
segundo comprende a todas las especies
actuales y extintas de vida terrestre,
incluidos dos géneros del Paleozoico (Compsoscorpius
y Palaeopisthacanthus). El trabajo
posterior de Stockwell (1989) y otros
análisis impiden aceptar la clasificación
propuesta por Kjellesvig-Waering (ver Tabla
nº 2 en la que se resumen ambas
clasificaciones y Anexo I, en el que se
relacionan todos los géneros de escorpiones
citados en este trabajo con ambas
clasificaciones). La clasificación y
filogenia propuestas por Stockwell (1989),
aunque con algunos problemas, ha sido
seguida por Selden (1993a) y Jeram (1994a,
1994b). Un esquema resumido se ilustra en la
Fig. nº 7. La Clase Scorpionida es dividida
en tres órdenes. Protoscorpiones
Petrunkevitch, 1949 y Palaeoscorpiones
Stockwell, 1989 serían grupos antiguos de
origen marino. El tercer orden (Scorpiones
Hemprich y Ehrenberg, 1829) estaría
compuesto por las restantes especies, todas
ellas terrestres. El primer representante
sería Praearcturus gigas Woodward,
1871 del Devónico, al que se unirían (en
cuanto a antigüedad) los restos de cutícula
canadienses de los mesoscorpiones estudiados
por Shear et al. (1996). El suborden
Mesoscorpionina Stockwell, 1989 incluiría
formas extintas con placas abdominales,
respiración aérea y una conformación de
coxa-esternón intermedia entre los
Palaeoscorpiones y las formas modernas. El
resto de especies se incluye bajo el
suborden Neoscorpionina (sensu Stockwell,
nom Kjellesvig-Waering) caracterizado por la
reducción del número de grupos de ojos
laterales, incremento de tricobotrias y
reducción de los dientes quelicerales. El
infraorden Paleosterni Stockwell 1989
incluye a especies del Carbonífero sin
estigmas en los esternitos ventrales,
constituyendo el grupo hermano de
Orthosterni Pocock, 1911 compuesto por los
géneros Palaeopisthacanthus (escorpión más
antiguo con estigmas respiratorios),
Compsoscorpius (con acusada reducción de los
ojos laterales y desplazamiento de los
medios) y las familias actuales (Stockwell,
1989; Jeram, 1994a, 1994b).
(1) Clasificación de Kjellesvig-Waering
(1986)
|
Orden: Scorpionida |
| |
|
Suborden: Branchioscorpionina |
| |
|
Infraorden: Holosternina
|
|
Infraorden: Meristosternina |
|
Infraorden: Lobosternina |
|
Infraorden: Bilobosternina |
| |
|
Suborden: Neoscorpionina |
| |
|
Infraorden: Orthosternina |
(2) Clasificación de Stockwell (1989 y
Selden (1993a):
|
Clase: Scorpionida
|
| |
|
Orden: Protoscorpiones |
|
Orden: Palaeoscorpioes
|
|
Orden: Scorpiones: |
| |
|
Suborden: Mesoscorpionina |
|
Suborden: Neoscorpionina |
| |
|
Infraorden: Palaeosterni
|
|
Infraorden: Orthosterni |
Tabla
nº 2: Clasificación de Scorpionida
según (1) Kjellesvig-Waering (1986) y
(2) Stockwell (1989) y Selden (1993a).
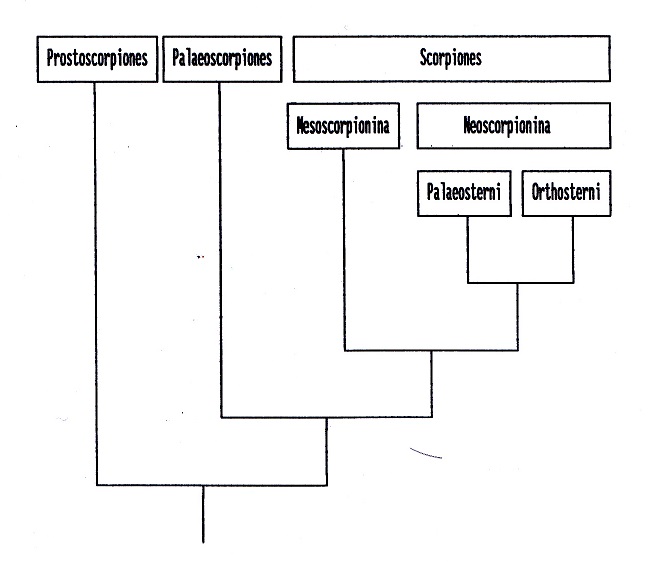
Figura 7:
Filogenia de Scorpionida propuesta por
Stockwell (1989) y Jeram (1994a, 1994b).
A pesar de los innumerables avances de la
última década, la clasificación y filogenia
de los escorpiones están todavía lejos de
una resolución satisfactoria. Entre el agua
y la tierra, el registro fósil de los
escorpiones nos permite reinterpretar
-aunque de momento con dificultades- la
larga historia geológica de un grupo de
artrópodos emblemático, pues, en definitiva,
el escorpión es, básicamente, un fragmento
del Paleozoico viviente.
Agradecimientos
A Enrique Peñalver por sus comentarios al
trabajo y su ayuda en la obtención de
bibliografía.
Notas:
(1) La forma
pentagonal es considerada como primitiva, ya
que aparece en los primeros estudios de
todas las especies (Pocock, 1894).
(2) Existen dos
importantes trabajos sobre sistemática de
los escorpiones fósiles pero de dificil
acceso: Stockwell (1989) y Jeram (1990a).
Uno de ellos, el segundo, de hecho, se
encuentra formalmente sin publicar.
(3) Estas
estructuras eran tan insospechadas que
Størmer en su descripción preliminar (1960)
supuso que se trataba de una invasión
parásita de hongos acaecida al individuo
después de su muerte y antes de su
fosilización.
(4) Otra
especie descrita es Tiphoscorpio hueberi
Kjellesvig-Waering 1986... que tras un
reciente estudio ha sido clasificado como un
"miriápodo" (Selden y Shear, 1992).
(5) Incluso en
un período anterior, el Ordovícico
(Kukalová-Peck 1991).
(6) El mismo
"truco" es utilizado por algunos crustáceos
mediante las llamadas cámaras branquiales.
(7) Stockwell
(1989) incluye en su clasificación el
Suborden Mesoscorpionina, al que también
asigna el género Mesophonus
(Triásico-Jurásico). Los restos del
escorpión del Devónico (en realidad, éste
sería el primer representante del grupo) y
los autores sugieren que todo el suborden
pudo ser terrestre.
(8) Dos
especies más han sido descritas de
Hispaniola T. quisqueyanus Armas y
T. crassimanus (Thorell), pero son
diferentes de las formas fósiles y
pertenecen a especie actuales (Poinar,
1992).
(9)
Especialmente si tenemos en cuenta que ambos
grupos son de origen marino y que ambos
participaron en la colonización del medio
terrestre. Scorpionida con indudable éxito;
Eurypterida mediante formas anfibias que,
como mínimo, eran capaces de realizar
prolongadas excursiones fuera del agua
(Selden, 1985; Rolfe, 1985; Shear y
Kukalová-Peck, 1990).
Bibliografía
ABD-EL-WAHAB,
A., 1952.-Some notes on the segmentation of
the scorpion, Buthus quinquestriatus
(H. & E.). Proceedings of the Egyptian
Academy of Sciences 7: 75-91.
ANDERSON, D.T.,
1973.-Embryology and phylogeny in
annedlids and arthropods. International
Series of Monographs in Pure and Applied
Biology, Zoology Division, vol. 50. Nueva
York, Pergamon, 495 pp.
BERGSTRÖM, J.,
1979.-Morphology of fossil arthropods as a
guide to phylogenetic relationships. En
GUPTA, A.P. (ed.): Arthropod Phylogeny.
Nueva York: 3-56.
BIRULA, A.A.B.,
1917.-Fauna of Russia and adjacent
countries. Arahnoidea, Vol.1,
Scorpions. Tr. B. Munitz. Jerusalem:
Israel Program for Scientific Translations,
1965, 154 pp. [citado en SISSOM, 1990]
BOUDREAUX, H.
B., 1979.-Arthropod phylogeny with
special reference to insects. Wiley,
Nueva York.
BRAUER, A.,
1985.-Beiträge zur Kenntnis der
Entwicklungsgeschichte des Skorpions, II.
Zeitschrift für wissenschaftliche Zoologie,
59: 351-433.
BROWNELL, P.H.,
1985.-Detección de la presa en el escorpión
de la arena. Investigación y Ciencia,
101: 52-60.
CAMPOS, D.R.B.,
1986.-Primeiro registro fóssil de
Scorpionoidea na Chapado do Araripe
(Cretáceo Inferior), Brasil. Anais da
Academia Brasileira de Ciências, 58:
135-137.
CARTHY, J.D.,
1966.-Fine structure and function of the
sensory pegs on the scorpion pecten.
Experientia, 22: 251-261.
CARTHY, J.D.,
1968.-The pectines of scorpions. Symposia
of the Zoological Society (London), 23:
251-261.
CLOUDSLEY-THOMPSON, J.L., 1955.-On the
function of the pectines of scorpions.
Annals and Magazine of Natural History,
ser. 12, 8: 556-560.
DUNLOP, J.,
1996.-Arácnidos fósiles (con exclusión de
arañas y escorpiones). Bol. SEA:
PaleoEntomología, 16.
DUFRE, G.,
1995.-Evolution historique du nombre de taxa
de l'ordre des Scorpionida. Revue
Arachnologique, 10(3): 223-227.
FOELIX, R.F. y
MÜLLER-VORHOLT, G., 1983.-The fine structure
of scorpion sensory organs. II, Pecten
sensilla. Bulletin of the British
Arachnological Society 6(2): 68-74.
FRANCKE, O.F.,
1982.-Are there any bothriurids (Arachnida,
Scorpiones) in southern Africa? Journal
of Arachnology, 10: 35-39.
FRANCKE, O.F. y
SOLEGLAD, M.E., 1981.-The family Iuridae
Thorell. Journal of Arachnology, 9:
233-258.
GANZELEWSKY, M.
y SLOTTA, R., 1996.-Bernstein tränen der
götter. Bochum.
HAMMEN, L. van
der, 1989.-An Introduction to Comparative
Arachnology. SPB Academic Pibl. bv, 576
pp.
HENRY, L.,
1949.-The nervous system and the
segmentation of the head in a scorpion
(Arachnida). Microentomology, 14(4):
121-126.
HJELLE, J.T.,
1990.-Anatomy and Morphology. En: POLIS, G.
(ed.): The biology of scorpions.
Stanford University Press, Stanford,
California: 9-63.
HONG, Y.,
1983.-Discovery of Miocene scorpius from the
diatoms of Shanwang in Shandong Province.
Bulletin Tianjin Institute, 8: 17-21 (en
chino).
HONG, Y., 1985.-Fossil
insects, scorpions and araneids in the
diatoms of Shanwang. Geological
Publishing House, Beijing, 80 pp.
JERAM, A.J.
1990a.-The Micropalaeontology of Paleozoic
Scorpions. Tesis no publicada, Univ.
Manchester [citado por Selden 1993a].
JERAM, A.J.
1990b.-Book-lings in a Lower Carboniferous
scorpion. Nature, 343: 360-361.
JERAM, A.J.,
1994a.-Scorpions from the Viséan of East
Kirkton, West Lothian, Scortland, with a
revision of the infraorder Mesoscorpionina.
Trans. R. Soc. Edinb., Earth Sc., 84:
283-288.
JERAN, A.J.,
1994b.-Carboniferous Orthosterni and their
relationship to living scorpions.
Palaeontology, 37(3): 513-550.
KJELLESVIG-WAERING, E.N., 1966.-Silurian
scorpions of New York. Journal of
Paleontology, 40(2): 359-375, 3 pl.
KJELLESVIG-WAERING, E.N., 1986.-A restudy
of the fossil Scorpionida of the World.
Paleontographica Americana, nº 55. Ithaca,
N.Y.: Paleontological Research Institute.
287 pp.
KUKALOVA-PECK,
J., 1991.-Fossil history and the evolution
of hexapod structures. En: CSIRO: The
Insects of Australia, I: 141-179.
LAMORAL, B.H.,
1980.-A reappraisal of suprageneric
classification of Recent scorpions and of
their zoogeography. En: Proceeding of the
8th International congress of arachnology,
Viena: 439-444.
LOURENÇO, W.R.,
1996.-The biogeography of scorpions. En:
Proceeding of the XIIIth International
Congress of Arachnology, Geneva. Revue
suisse de Zoologie, vol. hors sér. II:
437-448.
MAURY, E.A.,
1980.-Usefulness of the hemispermatophore in
the systematics of the scorpion family
Bothriuridae. En: Proceeding of the 8th
International congress of arachnology,
Viena: 335-339.
MELLO-LEITAO,
C., 1945.-Escorpioes sul-americanos.
Arquivos do Museu Nacional Río de Janeiro,
40, 468 pp.
MILLOT, J. y
VACHON, M., 1949.-Ordre des Scorpions. En:
GRASSE, P.P. (dir.): Traité de Zoologie.
Masson & Cie, París: 386-436.
MONZON, F. y
BLASCO, R., 1995.-In cauda venenum: El Mito
del Escorpión I: El escorpión en la
mitología y las leyendas. Bol. SEA,,
12: 33-38.
MONZON, F. y
BLASCO, R., 1996a.-In cauda venenum: El Mito
del Escorpión II: Simbolismo y bestiarios.
Bol. SEA, 13: 43-45.
MONZON, F. y
BLASCO, R., 1996b.-In cauda venenum: El Mito
del Escorpión III: Etimología de los
vocablos alacrán y escorpión. Bol. SEA,
14: 36.
MONZON, F. y
BLASCO, R., 1996c.-In cauda venenum: El Mito
del Escorpión IV: El escorpión en el
refranero. Bol. SEA, 15: 53-54.
PETRUNKEVITCH,
A., 1949.-A study of Palaeozoic Arachnida.
Transactions of the Conneticut Academy of
Arts and Sciences, 37: 69-315.
PETRUNKEVITCH,
A., 1953.-Palaeozoic and Mesozoic Arachnida
of Europe. Geological Society of America,
Memoir, 53: xi + 1-28 pp.
PETRUNKEVITHC,
A., 1955.-Arachnida. En MOORE, R.C. (ed.):
Treatise on Invertebrate Paleontology,
Part P, Arthropoda 2, Geological Society of
America & Univ. of Kansas Press: 42-162.
POCOCK, R.I.,
1894.-Scorpions and their geographical
distribution. Natural Science, 4(24):
353-364.
POCOCK, R.I.,
1901.-The scottish silurian Scorpions.
Quart. Microsc. Journal Sc., (2) 44:
291-311 + pl.19.
POCOCK, R.I.,
1911.-A monograph of terrestrial
Carboniferous Arachnida of Great Britain.
Palaeontograph. Soc. [1910], Londres, 84
pp.
POINAR, G.O.,
1992.-Life in Amber. Standord
University Press, Stanford, 350 pp.
POLIS, G.A.,
1990.-The Biology of Scorpions.
Stanford University Press, Stanford, 588 pp.
POLIS, G.A. y
SISSOM, D., 1990.-Life History. En POLIS,
G.A. (ed.): The Biology of Scorpions.
Stanford University Press, Stanford:
161-223.
ROBINSON, R.A.,
1987.-Chelicerata. En: BOARDMAN, R.S. et
al.: Fossil Invertebrates. Blackwell
Scientific Publ., Oxford: 258-264.
ROLFE, W.D.I.,
1985.-Early terrestrial arthropods: a
fragmentary record. Phil. Trans. R. Soc.
Lond., B 309: 207-218.
ROLFE, W.D.I. y
BECKETT, C.M., 1984.-Autoecology of Silurian
Xiphosurida, Scorpionida, and Phylocarida.
En: BASSET, M.G. y LAWSON, J.D. (eds.):
Autecology of Silurian Organisms. Special
Papers in Palaeontology, 32: 27-37.
SANTIAGO-BLAY, J.A. y POINAR Jr., G.O.,
1988.-A fossil scorpion Tityus geratus
new species (Scorpiones: Buthidae) from
dominican amber. Historical Biology,
1: 345-354.
SANTIAGO-BLAY, J.A. y POINAR Jr., G.O.,
1993.-First scorpion (Buthidae:
Centruroides) from mexican amber (Lower
Miocene to Upper Oligocene). The Journal
of Arachnology, 21: 147-151.
SANTIAGO-BLAY, J.A., SCHAWALLER, W. y POINAR
Jr., G.O., 1990.-A new specimen of
Microtityus ambarensis (Scorpiones,
Buthidae), fossil from Hispaniola: evidence
of taxonomic status and possible
biogeographic implications. The Journal
of Arachnology, 18: 115-117.
SAVORY, T., 1977.-Arachnida. 2d. ed.
Academic Press, London, 340 pp.
SCHAWALLER, W., 1979.-Erstnachweis eines
Skorpions in Dominikanischen Bernstein
(Stuttgarter Bernsteinsammlung: Arachnida,
Scorpionida). Stuttgarter Beiträge zur
Naturkunde, B 45: 1-15.
SCHAWALLER, W., 1982.-Zwei weitere Skorpione
in Dominikanischen Berstein (Stuttgarter
Bernsteinsammlung: Arachnida, Scorpionida).
Stuttgarter Beiträge zur Naturkunde,
B 82: 1-14.
SELDEN, P.A., 1985.-Eurypterid respiration.
Phil. Trans. R. Soc. London, B 309:
219-226.
SELDEN, P.A., 1990.-Fossil history of the
arachnids. Newl. Brit. Arachnol. Soc.,
58: 4-6.
SELDEN, P.A., 1993a.-Arthropoda
(Aglaspidida, Pycnogonida and Chelicerata).
En BENTON, M.A. (ed.) The Fossil Record
2.Chapman & Hall, Londres: 297-320.
SELDEN, P.A., 1993b.-Fossil arachnids-recent
advances and future prospects. Memoirs of
the Queensland Museum 33(2): 389-400.
SELDEN, P.A. y JERAM, A.,
1989.-Palaeophysiology of terrestrialization
in the Chelicerata. Trans. R. Soc. Edinb.,
80: 303-310.
SELDEN, P.A. y SHEAR, W.A., 1992.-A miriapod
identity for the Devonian "scorpion"
Tiphoscorpio hueberi. Bericht der
naturwissenschaftlich-Medezinischen Vereins
in Innsbruck, Suppl. 10: 35-35.
SHEAR, W.A. y KUKALOVA-PECK, J., 1990.-The
ecology of Paleozoic terrestrial arthropods:
the fossil evidence. Can. J. Zool.,
68: 1807-1834.
SHEAR, W.A., GENSEL, P.G. y JERAM, A.J.,
1996.-Fossils of large terrestrial
arthropods from the Lower Devonian of
Canada. Nature, 384: 555-557.
SHULTZ, J.W., 1989.-Morphology of locomotor
appendages in Arachnida: evolutionary trends
and phylogenetic implications. Zool. J.
Linn. Soc., 97: 1-56.
SISSOM, W.D., 1990.-Systematics,
Biogeography, and Paleontology, En: POLIS,
G. (ed.): The biology of scorpions.
Stanford University Press, Stanford,
California: 65-160.
STOCKMANN, R., 1991.-Les Scorpions.
Vie sauvage, 41, 20 pp.
STOCKWELL, S.A., 1989.-Revisión of the
Phylogeny and Higher Classification of
Scorpions (Clelicerata), Tesis no publicada.
Puede obtenerse copia microfilmada en Univ.
California, Berkeley.
STøRMER, L., 1960.-A
scorpion-like eurypterid from the Lower
Devonian of Germany. Internat. paleont.
Union, Proc. 21st Session Rept. Copenhagen,
pt. 22: 87-91.
STøRMER, L.,
1963.-Gigantoscorpio willsi, a new scorpion
from the Lower Carboniferous of Scartland
and its associated preying microorganisms.
Skr. norske. Vid.-Akad., Oslo, I,
mat.-naturvid. Kl, N.S., 8: 1-171.
STøRMER, L.,
1970.-Arthropods from the Lower Devonian
(Lower Emsian) of Alken an der Mosel,
Germany. Part 1: Arachnida.
Senckenbergiana lethaea, 51(4): 335-369.
STøRMER, L.,
1976.-Arthropods from the Lower Devonian
(Lower Emsian) of Alken an der Mosel,
Germany. Part 5: Myriapoda and additional
forms, with general remarks on fauna and
problems regarding invasion of land by
arthropods. Senckenbergiana lethaea,
57(2/3): 87-183.
VOGEL, B.R. y DURDEN,
C.J., 1966.-The occurrence of stigmata in a
Carboniferous scorpion. Journal of
Paleontology, 40: 655-658.
WEYGOLDT, P. y PAULUS,
H., 1979.-Untersuchungen zur Morphologie,
Taxonomie und Phylogenie der Chelicerata. I.
Morphologische Untersuchungen. II.
Cladogramme und die Entfaltung der
Chelicerata. Zeitschrift für zoologische
Systematik und Evolutionsforschung,
17(3): 85-116 + 177-200.
WILLS, L.J., 1947.-A
monograph of British Triassic scorpions.
Palaeontological Society, 100/101:
1-137, Londres.
WILLS, L.J., 1959.-The
external anatomy of some Carboniferous
"scorpions", part 1. Palaeontology
1(4): 261-282.
WILLS, L.J., 1960.-The
external anatomy of some Carboniferous
"scorpions", part 2. Palaeontology
3(3): 276-232.
|
ANEJO I |
| |
|
Relación
de géneros de escorpiones citados en
el artículo y clasificación según
diferentes autores. |
|
|
|
GENERO |
Clasificación según |
Clasificación según |
| |
Kjellesvig-Waering (1986 |
Stockwell (1989) + Jeram (1994a, b) |
|
________________________________________________________________________________________________ |
| |
|
|
|
Acanthoscorpio |
Branch.:
Holosternina |
incertae
sedis |
|
Allobuthiscorpius |
Branch.:
Holosternina |
Scorpiones: Neoscorpionina:
Palaeosterni |
|
Araripescorpio |
Neosco.:
Orthosternina |
Scorpiones: Neoscorpionina:
Orthosterni |
|
Archaeoctonus |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Archaeophonus |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Branchioscorpio |
Branch.:
Bilosternina |
incertae sedis |
|
Bromsgroviscorpio |
Branch.:
Lobosternina? |
? |
|
Brontoscorpio |
Branch.:
Holocternina |
? |
|
Buthiscorpius |
Branch.:
Holocternina |
Scorpiones: Neoscorpionina:
Palaeosterni |
|
Centruroides |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Compsoscorpius |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthosterni |
|
Dolichophonus |
Branch.:
Bilosternina |
Protoscorpiones |
|
Eobuthus |
Branch.:
Lobosternina |
Scorpiones: Mesoscorpionina |
|
Eoscorpius |
Branch.:
Lobosternina |
? |
|
Feistmantelia |
Branch.:
Lobosternina? |
? |
|
Gigantoscorpio |
Branch.:
Holocternina |
incertae sedis |
|
Hygroscorpio |
Branch.:
Holocternina |
? |
|
Isobuthus |
Branch.:
Lobosternina |
Scorpiones: Mesoscorpionina |
|
Liossoscorpiones |
Branch.:
Holocternina |
incertae sedis |
|
Mazonia |
Branch.:
Holocternina |
Scorpiones: Mesoscorpionina |
|
Mesophonus |
Branch.:
Holocternina |
Scorpiones: Mesoscorpionina |
|
Microtityus |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Mioscorpio |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Paläeolychas |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Palaeopisthacanthus |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Palaeophonus |
Branch.:
Lobosternina |
Protoscorpiones |
|
Palaeoscorpius |
Branch.:
Holocternina |
Protoscorpiones |
|
Phoxiscorpio |
Branch.:
Holocternina |
Scorpiones: Mesoscorpionina |
|
Praearcturus |
Branch.:
Holocternina |
incertae sedis |
|
Proscorpius |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Pseudoarchaeoctonus |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Scorpio |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Sinoscorpio |
Neosco.: Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Spongiophonus |
Branch.:
Holocternina |
incertae sedis |
|
Stenoscorpio |
Branch.:
Holocternina |
incertae sedis |
|
Stoermeroscorpio |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Titanoscorpio |
incertae sedis |
incertae sedis |
|
Tityus |
Neosco.:
Orthosternina |
Scorpiones: Neoscorpionina:
Orthostherni |
|
Waeringoscorpio |
Branch.:
Holocternina |
Palaeoscorpiones |
|
Willsiscorpio |
Branch.:
Holocternina |
incertae sedis |
Texto original:
Melic, A. 1996. El registro fosil de los
Escorpiones: entre el agua y la tierra. 16,
93-104. ver archivo completo en
PDF
________________________________________________________________________
Complementariamente puedes consultar: